{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

vel Cannopylea, vel Pansolenia (Pls. 99-128).
Phaeodaria, Haeckel, 1879.
Tripylea, Hertwig, 1879.
Cannopylea, Haeckel, 1881.
Pansolenia, Haeckel, 1878.
Definition.Radiolaria with a double membrane surrounding the central capsule, which bears on one pole of the main axis a peculiar astropyle, or a tubular main-opening, in the centre of a circular radiate operculum. Usually (but not constantly) a pair of small, lateral, accessory openings (or parapylae) on the opposite pole of the main axis. Extracapsulum constantly with a phaeodium, or with a voluminous aggregation of peculiar dark pigment bodies (phaeodella) covering the astropyle of the central capsule. Skeleton siliceous or silicated, always extracapsular, very rarely wanting. Fundamental form very variable, originally monaxon, often dipleuric or bilateral.
The legion PHAEODARIA, or CANNOPYLEA, in the extent here defined, was constituted by me in 1878, in my Protistenreich (p. 102) under the name PANSOLENIA. This name was given on the supposition that the skeleton of these interesting Radiolaria is always composed of hollow tubules, in contrast to that of the other Radiolaria, where it is never tubular. But I was soon convinced that this supposition was erroneous, that in a great part of the PANSOLENIA the skeleton is not composed of hollow tubules but of solid bars, and that a constant, very characteristic, and never failing mark of this group is to be found in the peculiar phaeodium, a voluminous, constant, extracapsular pigment body. Therefore, in 1879, I changed the name into PHAEODARIA, and having discovered in the collection of the Challenger an astonishing number of new and wonderful types of this group, I described, in a preliminary note on it, four different orders and ten families with thirty-eight genera (Ueber die Phaeodarien, eine neue Gruppe kieselschaliger mariner Rhizopoden; in Sitzungsberichte der Jenaischen Gesellschaft für Medicin and Naturwissenschaft. Sitzung vom, 12th December 1879).
In the same year (1879), Richard Hertwig, in his excellent work entitled Der Organismus der Radiolarien, published the first accurate description of the intimate structure of the soft body of the PANSOLENIA, and mainly of their central capsule; and having always observed, in the few representatives examined by him, three openings in the capsule (one main-opening and two accessory openings), he called them TRIPYLEA (loc. cit. p. 87), being guided by the erroneous supposition that these three openings are constant in all members of the group. But this is by no means the case. The two accessory openings are completely absent in several families, whilst in others their number is increased. A constant and very striking character, however, of all PHAEODARIA, is the peculiar structure of their tubular main-opening, which I call astropyle, with its radiate operculum and cannular proboscis. On account of this important and startling characteristic I proposed in 1881 to call this group CANNOPYLEA. The two names PHAEODARIA and CANNOPYLEA both express a very striking and quite constant character of these curious Radiolaria, whilst the two names PANSOLENIA and TRIPYLEA are applicable only to a part of the whole legion.
The history of our knowledge of the PHAEODARIA is short, but very remarkable. Although hundreds of species, many of them cosmopolitan, are distributed over all oceans and all zones, although their size is in general much greater than that of the other Radiolaria (usually 1 to 2 mm., often even 5 to 10 mm. or more), and although their form and structure are usually striking, nevertheless the PHAEODARIA remained completely unknown up to the year 1859. During that year I observed the first forms living in the Gulf of Messina, and described and figured in 1862 in my Monograph five genera and seven species, viz., (1) Aulacantha scolymantha (p. 263), (2) Thalassoplancta cavispicula (p. 261, now Cannobelos cavispicula), (3) Aulosphaera trigonopa, and Aulosphaera elegantissima (p. 359), (4) Spongodictyum trigonizon (p. 459, now Sagoplegma trigonizon), and (5) Coelodendrum ramosissimum, and Coelodendrum gracillirnum (p. 361). I recognised the structure of the three genera enumerated as 1, 3, and 5, as so remarkable and so different from that of the other Radiolaria, that I founded three peculiar families for them, the Aulacanthida, Aulosphaerida, and Coelodendrida.
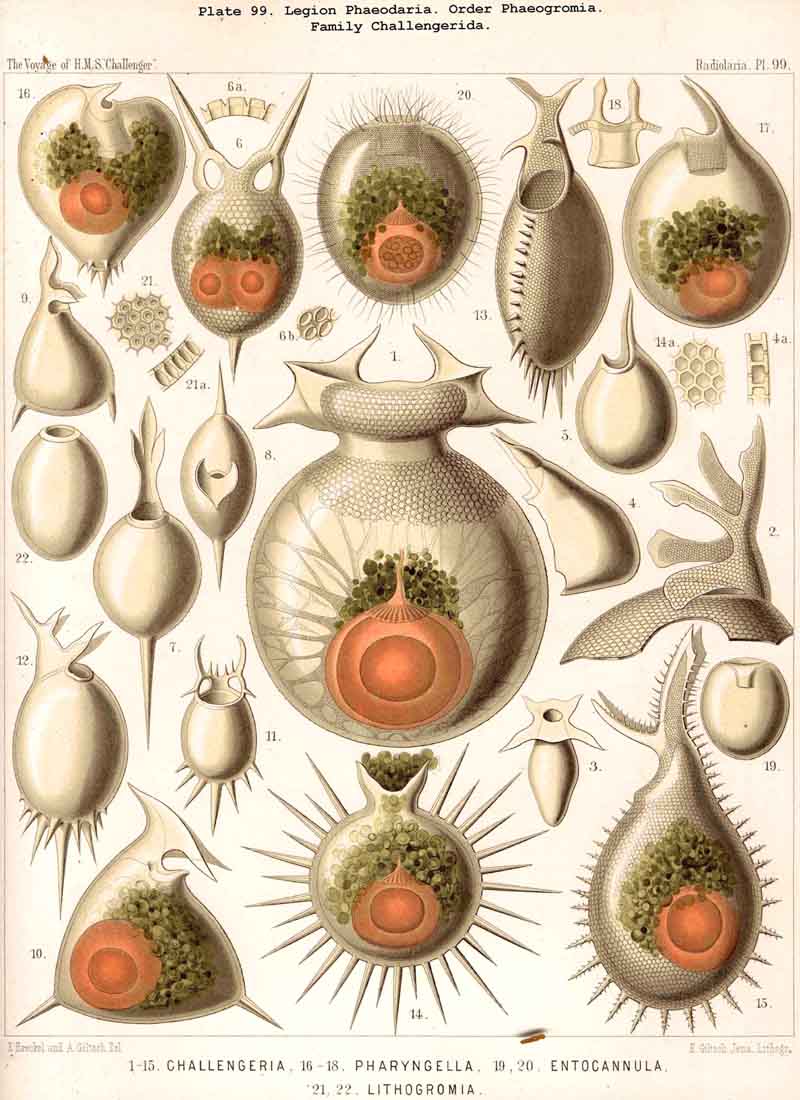
The first note on the numerous remarkable PHAEODARIA discovered by the Challenger, and mainly on the large-sized inhabitants of the deep-sea, was published in 1876 by Dr. John Murray, in his Preliminary Reports on Work done on Board the Challenger (Proc. Roy. Soc., vol. xxiv., read March 16, 1876). He pointed out (loc. cit., p. 535), that the tow-nets, sent down to a great depth (according to a new plan, adopted in April 1875) brought up on every occasion a great many new and pecnliar Rhizopods, which had never been observed in the nets used near the surface. "The shells of all have an exceedingly beautiful tracery, a fenestrated appearance often, which a closer examination shows to be caused by pit-like depressions. Some have only one, others have several openings, through which the sarcode flows. The sarcode of all these deep-sea Rhizopods has many large black-brown pigment-cells. At times they come up with a good deal of the sarcode outside of the shell; and two specimens have been seen to throw out elongated pseudopodia" (loc. cit., p. 536). Dr. John Murray distinguished at that time not less than fifty species of these interesting deep-sea Rhizopods and called them provisionally Challengerida; a term which we retain here for the largest and most characteristic family. He gave at the same time, in an accompanying plate (xxiv.), six figures of new PHAEODARIA, the names of which (as found by me in 1879 in the corres ponding preparations) are the following; (1) Challengeria naresii, (2) Challengeria aldrichii, (3) Bivalva compressa (now = Conchopsis compressa), (4) Tuscarora belknapii, (5) Challengeria circopora (now = Circoporus sexfurcus), and (6) Haeckeliana porcellana. A great number of these Challengerida (twenty species) were afterwards figured by Dr. John Murray in the Narrative of the Cruise of H.M. S. Challenger, 1885, vol. i. part 1, p. 226, Pl.A; viz., fourteen species of Challengeria and six species of Tuscarora.
The most important advance in the knowledge which we had of the peculiar organisation of the PHAEODARIA, was made by the accurate description which Richard Hertwig published, in 1879, of the intimate structure of their central capsule, and mainly of its peculiar openings. He examined living at Messina the following three forms, described in my Monograph; (1) Aulacantha scolymantha, (2) Aulosphaera elegantissima, and (3) Coelodendrum ramosissimum. Besides, he described an interesting new genus, Coelacantha anchorata; and another new form, which he placed in the Aulosphaerida, as Aulosphaera gracilis, but which really was a new genus of Sagosphaerida, here described as Sagoscena gracilis. Finally, Hertwig first discovered that the peculiar bodies, described by Ehrenberg as Dictyocha and placed by him in the Diatomea (Polygastrica), were the isolated pieces of the skeleton of a true Phaeodarium, and that they were scattered loosely in great numbers over the surface of the jelly-sphere, just as are the hollow spicula of Thalassoplancta or Cannobebos.
The six species mentioned, of which Hertwig gave a very accurate description and very instructive figures, belong to six different genera, and these represent six different families of PHAEODARIA, viz.; Aulacanthida, Aulosphaerida, Coelodendrida, Cannosphaerida, Sagosphaerida and Cannorrhaphida. He found that all these six forms, in spite of great differences in the form and structure of their skeleton, were identical in the structure of the central capsule; and since he observed constantly three openings in its double wall (a large main-opening on the oral pole, and a pair of lateral accessory openings on the aboral pole of its main axis) he called them TRIPYLEA (loc. cit., p. 87, 94). But he also pointed out the remarkable shape of their voluminous extracapsular body, and especially the characteristic position, size, colour and composition of the large pigment body, which I had called the phaeodium.
The accurate description of the gigantic and elegant skeleton of a new Phaeodarium, surpassing all other known Radiolaria in its extraordinary size (15 mm.), was published in 1882 by O. Bütschli (in Zeitschr. f. wiss. Zool., vol. xxxvi. p. 486, Taf. xxxi.). He called it Coelothamnus davidoffii, in honour of its discoverer, who had found it floating on the surface of the Gulf of Villafranca, near Nice. He placed it among the Coelodendrida; it belongs, however, to that part of this group which possesses a nasal tube, and which I afterwards separated under the name Coelographida.
The total number of PHAEODARIA, hitherto described and illustrated by figures, amounts therefore to seventeen species, viz., the seven species first described by myself (in 1862); the six new species figured by Dr. John Murray (in 1876); the three new species discovered by Hertwig (in 1879) and the single species last-mentioned described by Bütschli (in 1882). The rich collection of the Challenger has added to this small number such an astonishing wealth of new and remarkable forms, that I can describe in the following system of PHAEODARIA not less than eighty-four genera and four hundred and sixty-five species. These belong to fifteen different families and four different orders. But this great number is probably only a small part of the numerous interesting PHAEODARIA, which are abundantly distributed over all the oceans; those (e.g.) of the Indian and of the Arctic Oceans are almost unknown.
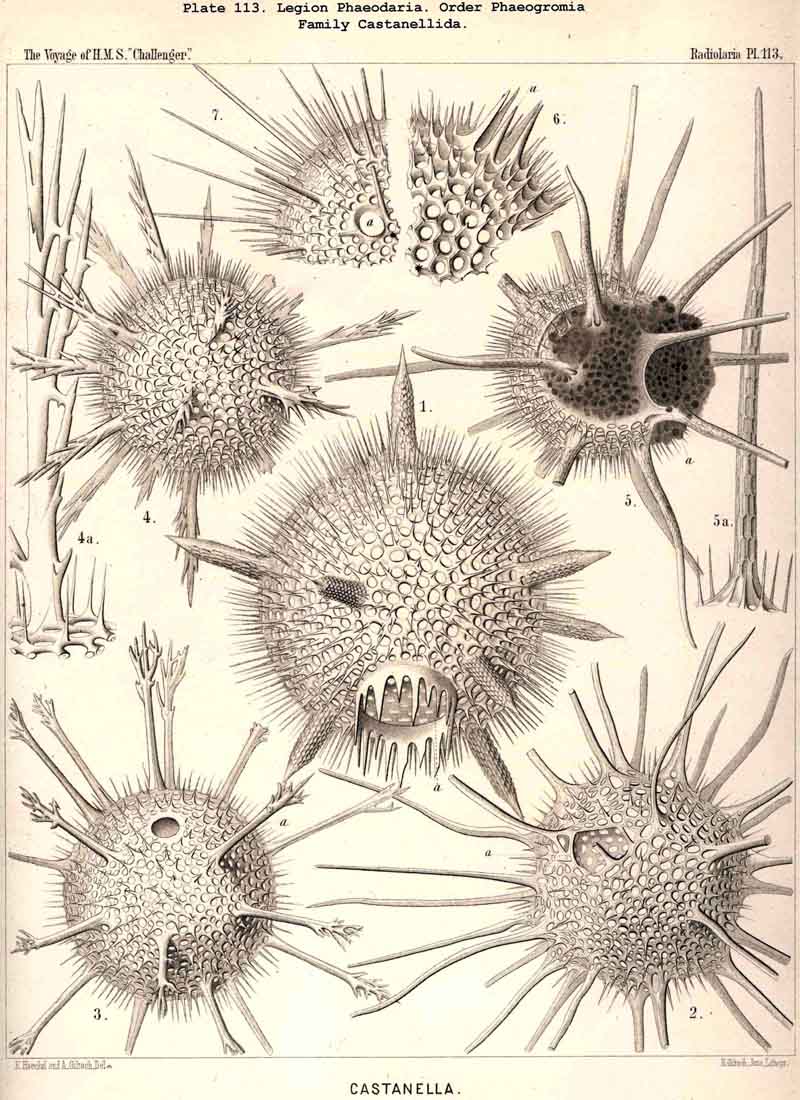
The great majority of these wonderful PHAEODARIA are inhabitants of the deep-sea, mainly of the southern hemisphere, and are so common in many stations explored by the Challenger, that its collection contains many thousands (or rather hundreds of thousands) of well-preserved specimens. A smaller part of the legion is found on the surface, widely distributed over all oceans; some of these are very common (as, e.g., Aulacantha, Aulosphaera, Sagosphaera, Coelodendrum, Castanella, &c.) and it is difficult to explain how they could entirely escape the eyes of all former observers.
The three general characters which distinguish the PHAEODARIA easily and constantly from all the other Radiolaria are the following :(1) the double membrane, a thick outer and a thin inner envelope, of the big central capsule; (2) its typical main-opening or astropyle, placed on the oral pole of the main axis, and distinguished by a peculiar radiate operculum, with tubular proboscis; (3) the phaeodium, or the peculiar voluminous pigment-body, which constantly lies in the oral half of the calymma, surrounds the oral part of the central capsule, and is composed of numerous phaeodella, or singular pigment-granules of green, olive, brown or black colour.
Besides these three general and never failing marks of the PHAEODARIA, the majority of this legion (but by no means all) possess the three following peculiarities; (1) two parapylae or accessory openings of the central capsule, placed laterally (at the right and left) on the aboral pole of the main axis (wanting in the Challengerida, Medusettida, Castanellida, and perhaps in some other families); (2) a characteristic skeleton which is always extracapsular, wanting only in the Phaeodinida, incomplete in the Cannorrhaphida and Aulacanthida, but perfectly developed and of very various shapes in the twelve other families; usually this silicated skeleton is composed of hollow tubules, which are filled up by jelly (PANSOLENIA); but in some families it is composed of ordinary solid network, not different from that of the other Radiolaria, e.g., especially in the Castanellida and Sagosphaerida; (3) an extraordinary size of the body, a.s well of the central capsule and its nucleus, as of the extracapsular skeleton; the majority of PHAEODARIA have a diameter of 1 to 2 mm., and are therefore from ten to twenty times as large as the majority of the other Radiolaria; in some gigantic forms the diameter of the unicellular body reaches 20 or even 30 mm.
The Central Capsule of the PHAEODARIA is the most important part of their body and preserves in all families of this legion the same essential structure, and nearly the same form, in striking contrast to the extraordinary variety and complication of the skeleton. As already mentioned, it differs from the central capsule of all the other Radiolaria in two most important peculiarities; firstly, the double membrane of the spheroidal capsule, and secondly, the singular structure of its constant main-opening, the astropyle. A third constant character is the considerable size of the enclosed nucleus, the diameter of which usually equals about half of that of the capsule itself.
The diameter of the capsule is, in the majority of PHAEODARIA, 0.1 to 0 .2 mm., often also 0.3 to 0.4, rarely more than 0.5, or less than 0.05 mm.
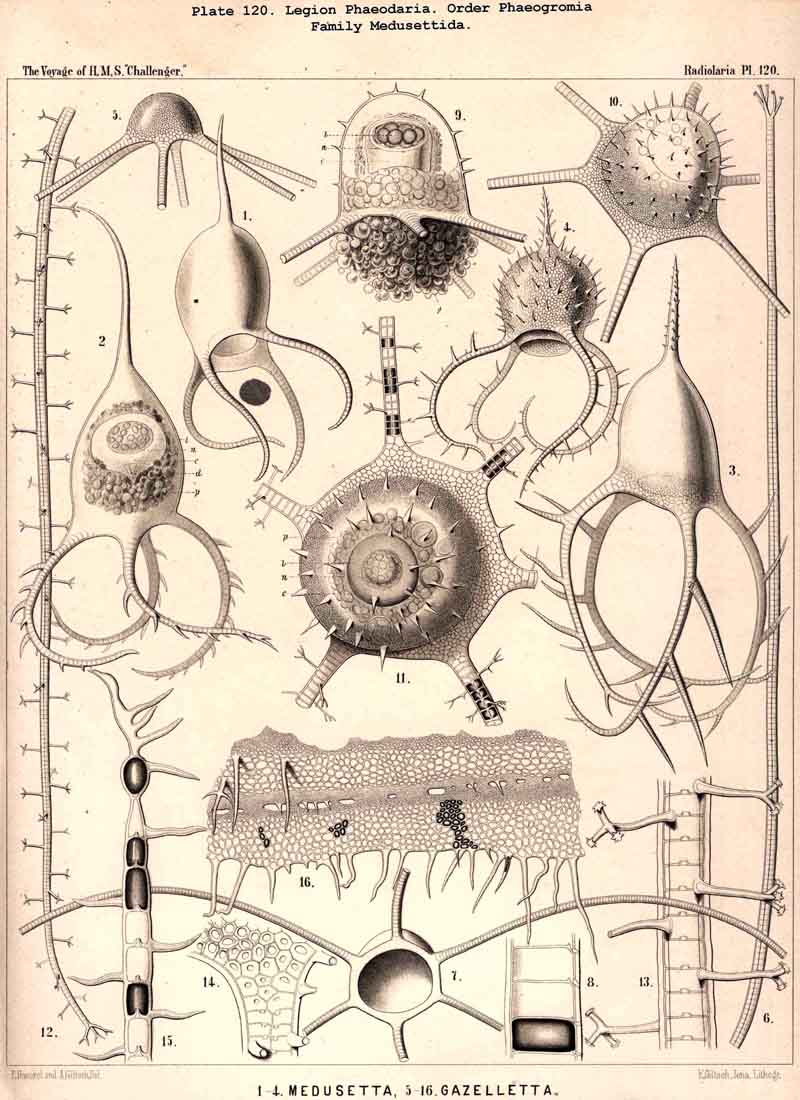
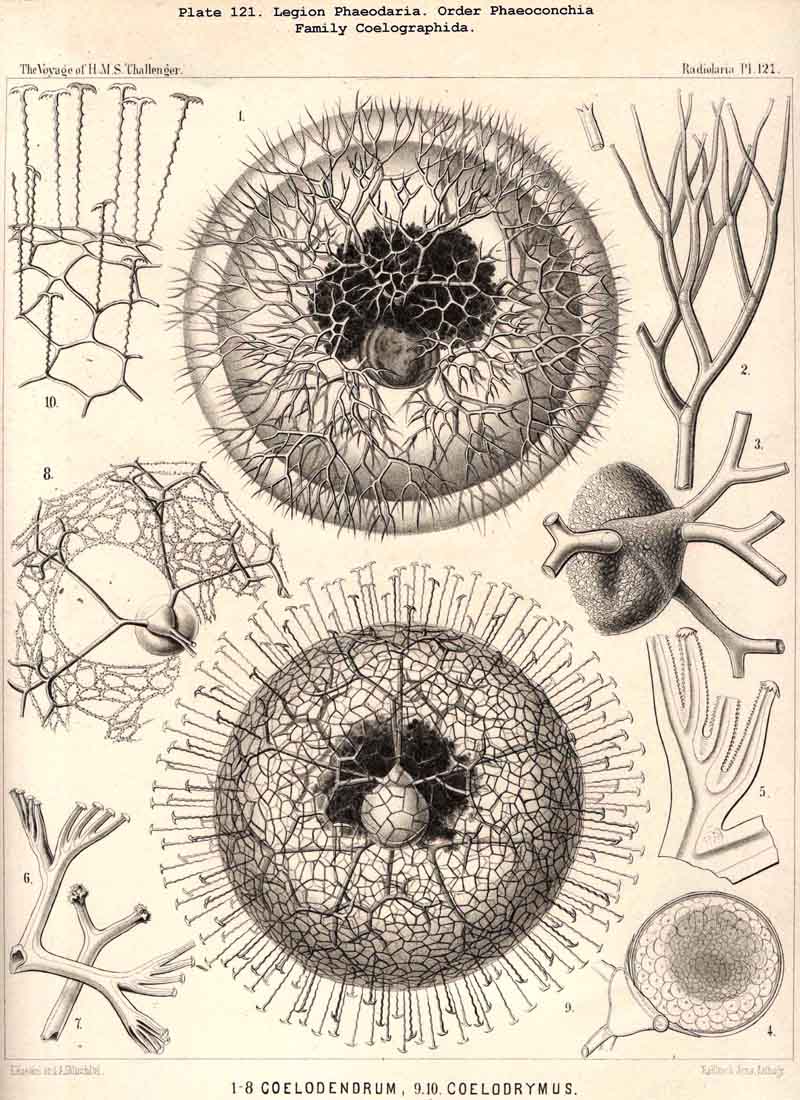
The position of the central capsule is somewhat different in the four orders into which we have divided the PHAEODARIA. It lies in the centre of the spherical, concentric calymma in all the Phaeocystina, or in those PHAEODARIA which possess no complete lattice-shellPhaeodinida, Cannorrhaphida, and Aulacanthida (Pls. 101 -105). The Phaeosphaerida (comprising the Orosphaerida, Sagosphaerida, Aulosphaerida, and Cannosphaerida, Pls. 106 - 106) all possess a very big, usually spherical lattice-shell, and here the central capsule is much smaller than the latter and lies in its centre. Another position is constantly occupied by the central capsule in all Phaeogromia (Pls. 99, 100, 113 -120; the families Challengerida, Medusettida, Castanellida, Circoporida, Tuscarorida), which have a peculiar mouth on the lattice-shell, placed on the oral pole of its main axis; the central capsule lies in the opposite aboral half of the shell cavity. The Phaeoconchia finally (Pls. 121 -128, the families Concharida, Coelodendrida, and Coelographida) all possess a bivalved shell, and the capsule is here enclosed between the two valves of the shell.
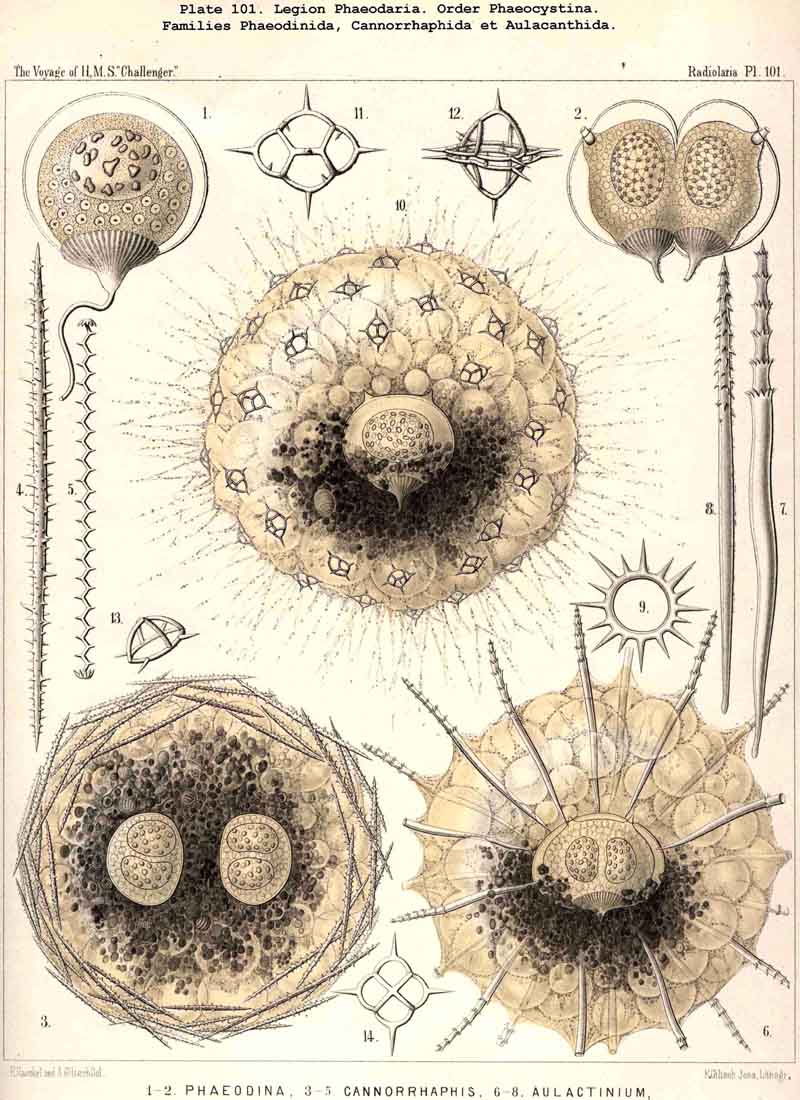
The form of the central capsule is constantly spheroidal, slightly depressed in the direction of the main axis, and therefore comparable to the spheroidal form of our globe. The depression is generally very slight, so that the proportion of the minor vertical main axis to the major horizontal or equatorial axis is about 4 : 5 or 5 : 6, often even 8 : 9 or less; but sometimes the proportion becomes 3 : 4, or even 2 : 3, so that the capsule becomes nearly lenticular; very rarely its form becomes almost perfectly spherical (Pl. 101, fig. 1). The main axis stands always vertically, and is distinctly marked by the peculiar shape of the astropyle, placed on its oral pole. Usually this pole, in the living and freely floating PHAEODARIA seems to be the lower pole, directed downwards (as also in the NASSELLARIA); but in some families it seems to be inversely directed upwards, as in the Challengerida and Tuscarorida (Pls. 99, 100).
The two membranes of the central capsule possess in all PHAEODARIA a very different shape, and were in all preparations which I could accurately examine (some hundreds belonging to all families) separated by a clear, rather wide interval, filled up either by a colourless fluid or by a structureless jelly (Pl.103, fig. 1; Pl.123, figs. 8, 9 ,&c.). They are in direct connection only at the openings. In the living PHAEODARIA, however, their distance is very small, or they are in immediate contact without any interval (Pl.101, fig. 10; Pl.102, fig. 1). According to the observations of Hertwig, the two membranes are always in close contact, and without interspace, in the living PHAEODARIA; and the space between them is an artificial product due to the influence of the preserving fluid or of certain chemical agents. In every case it is very easy to separate both membranes completely, except at the openings, where they are in direct connection. We distinguish both membranes shortly as ectocapsa and endocapsa.
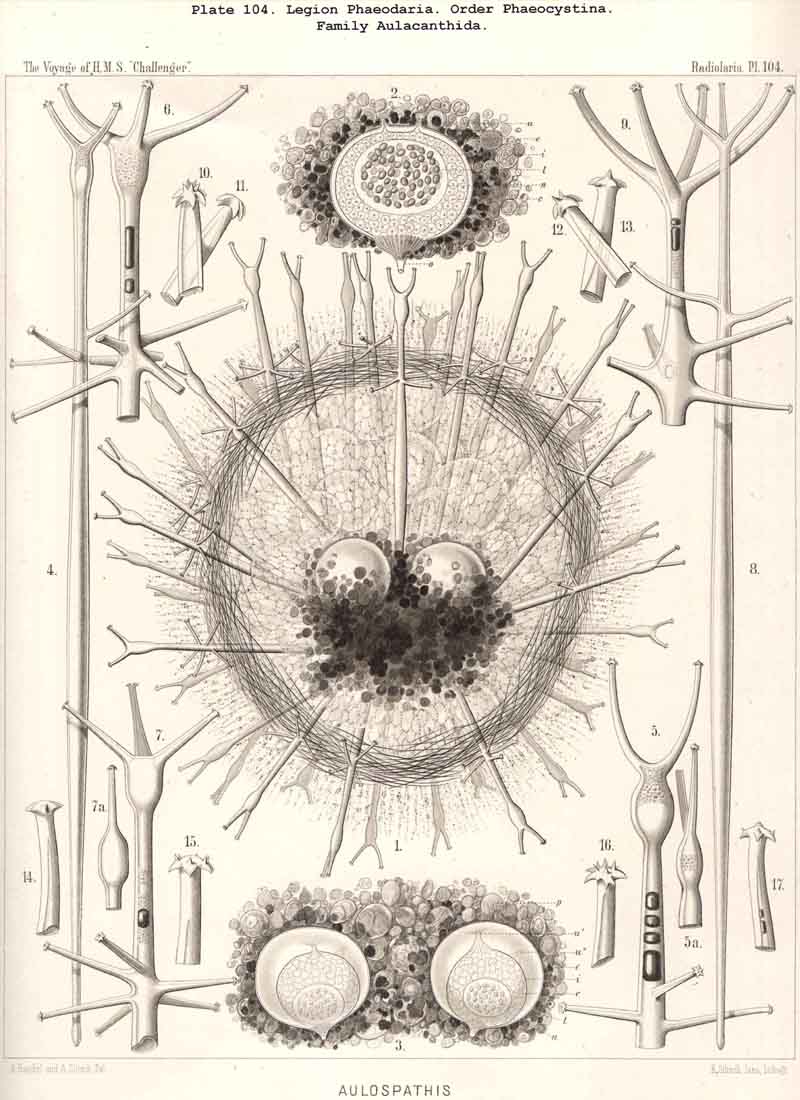
The ectocapsa, or the outer membrane of the central capsule, is rather firm and durable, double-contoured, elastic and difficult to destroy. Its physical and chemical qualities seem to approach those of chitin. It becomes, however, stained red by carmine, and yellow by nitric acid. Usually it appears structureless and refracts the light strongly. In a few cases, however, it exhibits, when examined by strong lenses, a fine punctation; and in some Aulacanthida (especially in some big forms of Aulographis and Aulospathis) the entire ectocapsa was densely covered with peculiar curved, or S-shaped dark corpuscles (Pl.114, fig. 13). They were all of the same length, about 0.01, and seemed to lie on its inner face.
The endocapsa, or the inner membrane of the central capsule, is much thinner than the outer, with which it is in immediate connection only at the openings. It encloses the entire contents of the capsule, and becomes very distinct, as soon as the latter are dissolved by chemical agents, or stained by carmine. In the majority of well-preserved preparations it is irregularly plicated, and resembles a thin, but firm, crumpled paper. Isolated pieces of the endocapsa are completely structureless, but exhibit also a considerable resistance, in spite of their minute thickness.
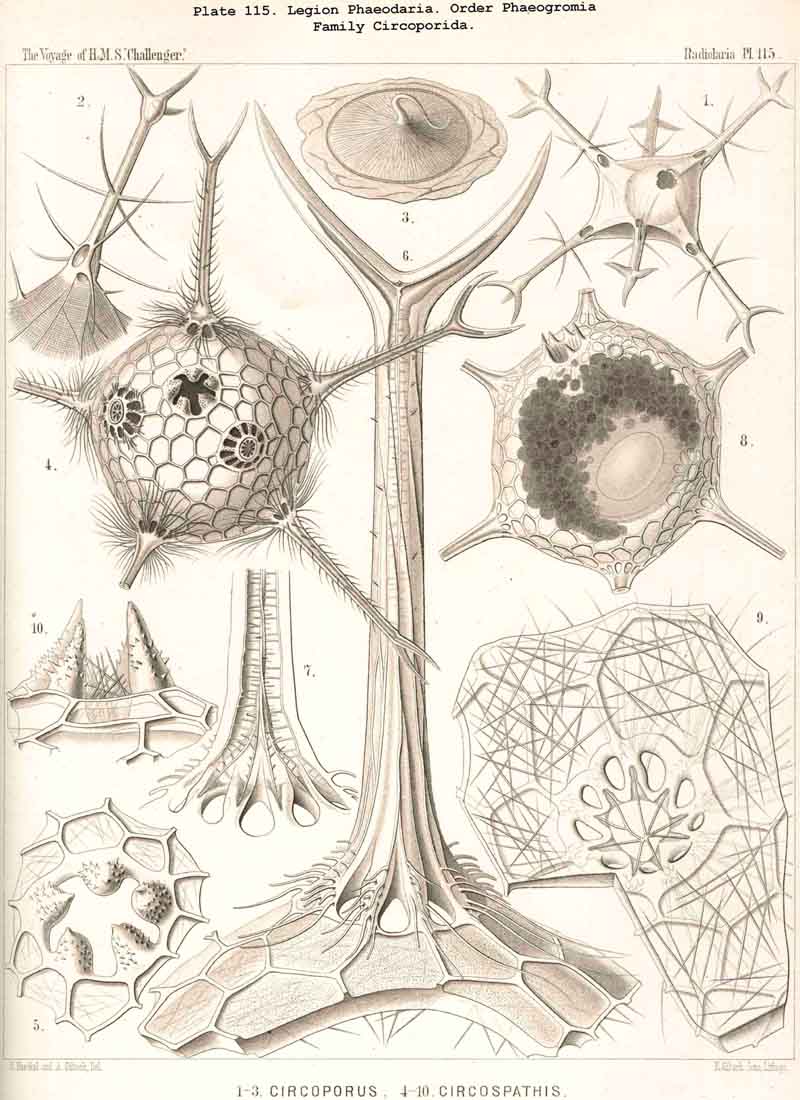
The openings of the central capsule exhibit in the PHAEODARIA a greater variety than their discoverer, R. Hertwig, supposed. The majority of the legion, certainly, possess the three openings described by him, and are therefore true TRIPYLEA. Some families, however, have only one opening, the astropyle, which is generally present (Challengerida, Medusettida, Castanellida, and Phaeocolla among the Phaeodinida, Pl. 101, fig. 1). In some other families there is a variable number of accessory openings or parapylae, one, three, or more, e.g., especially in the Circoporida and Tuscarorida. The former may be called Astropylea, the latter Sporopylea. Thus only a single opening to the central capsule is constant in all PHAEODARIA without exception, and that is the astropyle, or the large main-opening with its peculiar structure.
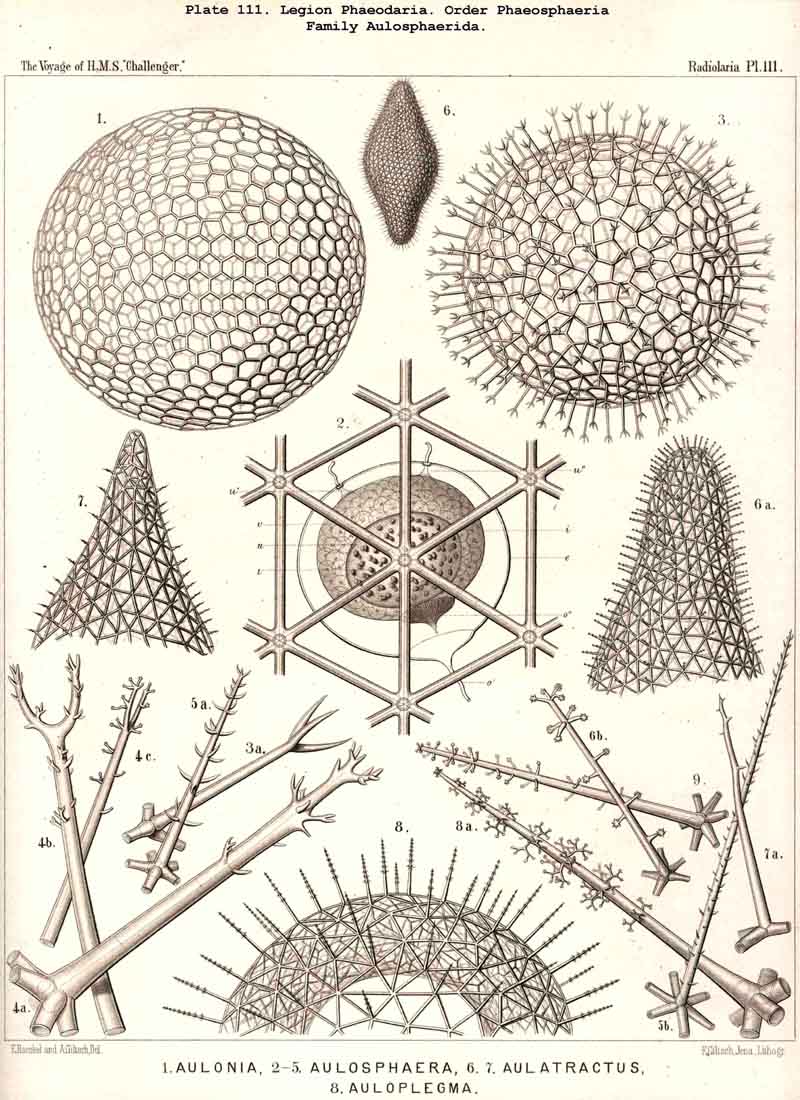
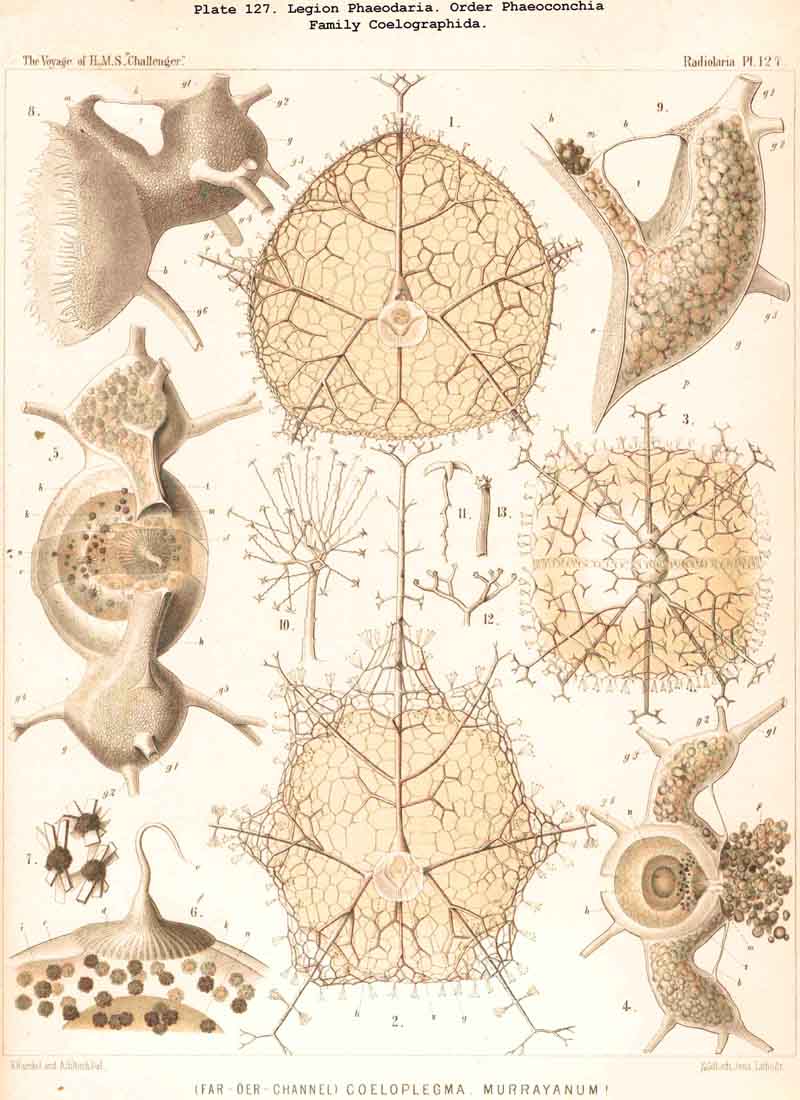
The astropyle, or the single constant main-opening of the central capsule, is distinguished by a very remarkable structure, and is sufficient of itself to separate the PHAEODARIA from all the other Radiolaria, and from the other Rhizopoda in general. It is always placed on the oral pole of the main axis, forming here a peculiar cap-shaped or flatly conical elevation, the centre of which is prolonged into a short, cylindrical tubule; we call the latter shortly a proboscis, and the former an operculum. To understand better the different forms which these important parts assume in the different families, compare Pl. 101, figs. 1-6; Pl. 102, fig. 1; Pl. 103, fig. 1; Pl. 104, figs. 1-3; Pl. 111, fig. 2; Pl. 123, figs. 1-9; Pl. 127, figs. 4-6; Pl. 128, fig. 2, &c. Compare also the first very accurate figures which R. Hertwig has given in 1879 (loc. cit.) in his Taf. x.
The operculum of the astropyle, the most important part of this main opening (" der Oeffnungshof der Hauptöffnung," in the description of Hertwig), is a circular convex plate, always more strongly vau'lted than the surrounding part of the capsule membrane, and is sharply separated from it by a circular, often thickened and double-contoured margin. The operculum covers the main-opening like the lid of a tea kettle, and the proboscis arising from its centre is comparable to the handle of that lid. The diameter of the circular operculum is usually about half as long (rarely as long) as the radius of the central capsule, therefore in the majority of PHAEODARIA 0.03 to 0.06, sometimes 0.1 and more. Its form is sometimes more conical, sometimes more like a mamma. Its height is usually about equal to half its diameter. It always exhibits a very distinct radial striation, produced by numerous prominent radial ribs, which arise in the centre and end at the circular sharply truncated margin. The usual number of these radial ribs may be, in the operculum of smaller capsules, from thirty to fifty, in those of medium size from sixty to ninety, and in the largest two hundred to three hundred or more. Usually the ribs are simple (Pl. 101, figs. 1, 2, 6, 10; Pl. 127, figs. 4-6; also in all figures given by Hertwig); but sometimes, mainly in the biggest Aulacanthida, they are distinctly branched in a centrifugal direction (Pl. 114, fig. 13; Pl. 115, fig. 3). Hertwig is of the opinion that the radial ribs are thickened ridges of the endocapsa ("leistenartige Verdickungen der inneren Membran"), and that the ectocapsa covering it is structureless (compare his figs. 1 and 1a, in Taf. x.). But I am convinced now, by numerous experiments and observations, that the radiate operculum is a part of the outer, not of the inner membrane. That part of the latter which lies immediately beyond the former, and which may be called the "inner operculum," also exhibits usually a fine radial striation; but this is probably only the cast of the stronger and much more prominent radial ribbing of the "outer operculum" belonging to the ectocapsa. The latter exhibits a large circular opening with thickened margin, when the radiate operculum is taken from it. The operculum becomes stained intensely red by carmine, yellow by nitric acid, corresponding to the ectocapsa, the main-opening of which is closed by it.
The proboscis, or the cylindrical tubule, which arises in the centre of the operculum, is often rather short, and bears the same relation to it as the nipple does to the human breast (Pl. 104, fig. 2). But usually it is more or less prolonged and often about as long as the radius of the capsule, rarely nearly as long as its diameter (Pl. 101, fig. 1). Its form is usually a slender cylinder, sometimes somewhat conical and tapering towards the distal end. It is more or less curved or S-shaped in the majority of preparations (Pl.115, fig. 3 Pl.127, fig. 6, &c.). Its thin wall is a direct prolongation of the outer membrane of the capsule, therefore it appears as a direct apophysis of the operculum, when this is detached from the latter. The opening at the distal end of the proboscis, through which the endosarc is thrown out, is circular.
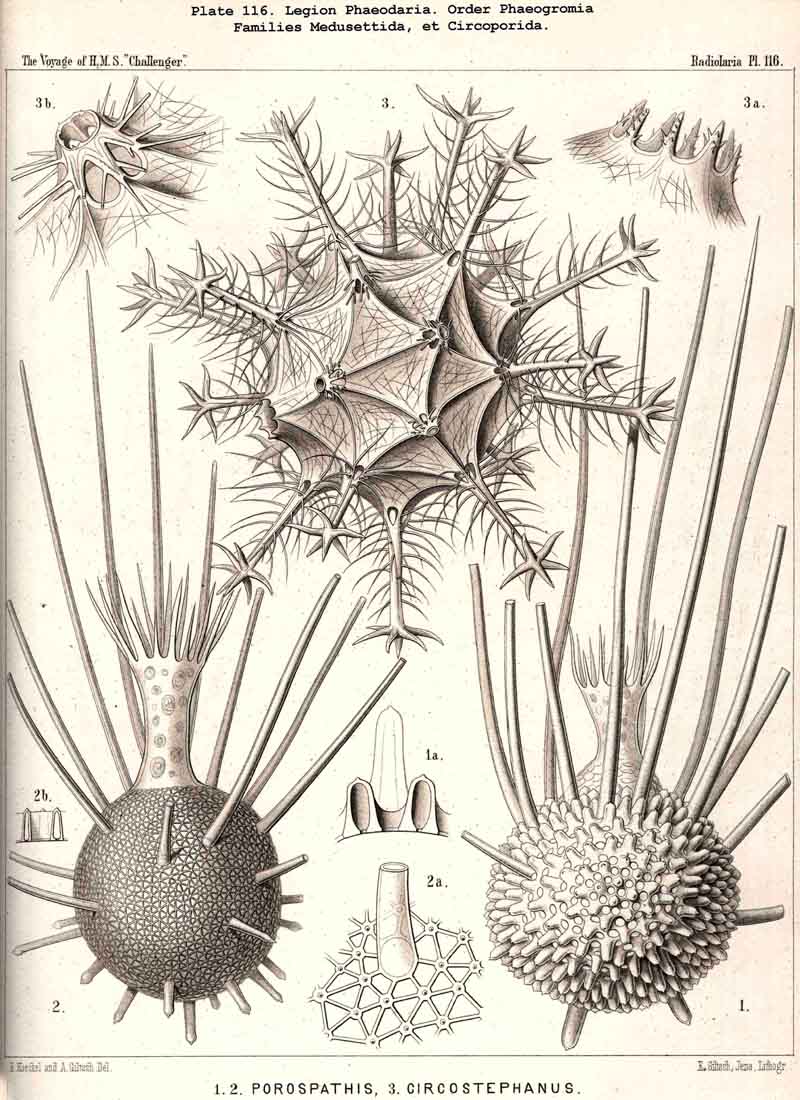
The parapylae, or the accessory openings of the central capsule, exhibit in all PHAEODARIA the same form and structure, but vary in number and disposition. They are recognised with difficulty in the smaller species, since they are always of small size, and may be easily overlooked. They seem to be completely wanting in the following families :-Challengerida (Pl.99), Medusettida (Pls. 118 -120), Castanellida (Pl. 113), and in single genera of other families, as in Phaeocolla (Pl.101, fig. 1). The majority of PHAEODARIA seem to possess the two lateral parapylae, first described by Hertwig, placed at a variable distance on each side of the aboral pole, to the right and left (Pl.101, figs. 2, 6, 10; Pl.103, fig. la; Pl.104, figs. 1, 2a; Pl. 123, figs. 1, 8a, &c.). The horizontal axis, in which the two parapylae lie, is the frontal or lateral axis; and the plane, which passes through the three openings of these "Tripylea," is the frontal or lateral plane. The number of the parapylae seems to be variable in the two families, Circoporida and Tuscarorida, which also differ from the other PHAEODARIA in the peculiar (porcellanous) structure of their shell-wall. The number seems to vary even in the single species of one genus; and the following cases may be found : (A) a single parapyle, placed on the aboral pole of the main axis and directly opposed to the astropyle (on the oral pole), as in Tuscaridium (Pl.100, fig. 8); (B) three equidistant parapylae, one of which is placed in the sagittal plane, and the two others one on each side of it, to the right and left, as in Tuscarora (Pl.100, figs. 1-6); the three parapylae seem to correspond here to the three long feet, or the tubular aboral apophyses of the shell, which are arranged in a similar manner to the three cortinar feet of the NASSELLARIA; (C) four equidistant parapylae, placed in pairs on the poles of the diagonal axes of a horizontal plane, as in Tuscarusa (Pl.100, fig. 7); they seem here to correspond to the four crossed aboral feet; (D) six to eight or more parapylae, probably in the different genera of Circoporida (Pls. 114 - 117); perhaps each radial tube, which is surrounded on its base by a circle of pores, here corresponds to a separate parapyle. The number of well-preserved central capsules belonging to the two latter families, however, which I could examine was relatively small, and their examination very difficult; therefore these numbers are not stated with satisfactory certainty and require a further more accurate examination. The minute structure of the parapylae (or "Neben-Oeffnungen ") has been described already very accurately by Hertwig (loc. cit., p. 95, Taf. x. figs. 1-11b). The two membranes of the capsule are in direct and immediate connection on each parapyle. The strong outer membrane arises in the form of a ring or of a short cylindrical tubule (collare paraboscidis or "Oeffnungshals "), is then reflected inwards, and connected at the bottom of the cylindrical cavity with the delicate inner membrane. From this connective ring arises a short conical or cylindrical tubule, which we call shortly the "paraboscis." According to Hertwig (who calls it "Oeffnungskegel ") the paraboscis is a direct prolongation of the inner membrane only. My own observations have led me to the opinion, that the paraboscis of each parapyle is a direct prolongation of the outer membrane (similarly to the larger proboscis of the astropyle), and that the basal connective ring is, therefore, the inner openi'ng of the paraboscis, through which the entosarc enters, being protruded through its outer circular opening. Usually the paraboscis of each parapyle is only a short cylinder, arising by a conical base; but sometimes, especially in some Aulosphaerida and Sagosphaerida, it is prolonged into a slender tubule, nearly as long as the radius of the central capsule. It has been already figured by Hertwig (loc. cit.,Taf. x. figs. 6-8). It seems, therefore, that the paraboscis of the accessory openings is developed in a way similar to the proboscis of the main-opening, and that the chief difference between the two is indicated by the large radiate operculum of the latter.
The cavity of the endocapsa, or the spheroidal space enclosed by the inner membrane of the central capsule, is filled up in its central part by the big nucleus, in its peripheral part by the endoplasm, or by the internal protoplasm, which is in communication with the outer or extracapsular protoplasm by the openings of that capsule. The endoplasm, or the intracapsular sarcode, is rather opaque, finely granulated, and usually filled up by numerous small clear spherules of equal size, which are more or less regularly arranged and equidistant. These spherules have usually a diameter of 0.01 to 0.015, rarely more than 0.02 or less than 0.005 mm.; their size is generally equal in each capsule. They have been already described and figured in my Monograph, as "wasserhelle kugelige Bläschen," and are probably vacuoles or small vesicles filled up by jelly or by a clear fluid. Usually each vesicle contains a small dark granule of fat, or a group of such granules connected together; and in these Hertwig observed a vibrating molecular motion. The central capsule of many PHAEODARIA contains, besides the vacuoles, often granules of pigment (usually red or yellow) and sometimes numerous groups of small crystals, placed mainly beyond the operculum of the astropyle (Pl.127, figs. 4-7). The outer layer of the endoplasm, placed immediately beyond the endocapsa, often exhibits a fine striation, as if composed of delicate fibrillae. This fibrillar striation is usually most distinct on the base of the openings, where also the endoplasm becomes stained very intensely by carmine. The astropyle as well as the parapylae often exhibit here, when observed from the face, a distinct radial striation (compare Hertwig, loc. cit., p. 98, Taf. x. figs. 6-14). Perhaps the radiate shape of the operculum is only produced by radial folds of the endocapsa placed beyond it, and these folds may be in turn the product of the radial fibrillae, which are prominent beyond the astropyle. On the other hand, these fibrillae may be compared to the muscular fibrillae or "myophane filaments" in the ectosarc of the Infusoria, and may perhaps effect by their contraction a dilatation of the openings of the capsule.
The nucleus of the PHAEODARIA is always very large, usually about half or two-thirds as broad as the central capsule, and placed either in its centre, or sometimes nearer to one pole of the main axis, which is common to the capsule and its nucleus. Therefore the diameter of the latter is usually half or even two-thirds of that of the capsule, and may be in the majority 0.05 to 0.15, often 0.2 to 0.3, rarely more than 0.4 or less than 0.01 mm. The form of the nucleus is rarely spherical, usually spheroidal, and it is more depressed in the direction of the main axis than the capsule itself. In my Monograph, where I gave the first description of it, I called it "Binnenbläschen." The membrane of the vesicular nucleus is thin but rather firm, and contains a rather clear, finely granulated substance, in which numerous nucleoli are usually scattered. (Compare Pl.101, figs. 1-10; Pl.102, 103, 104; Pl.123; Pl.127, &c.; the nucleus is marked by n., the nucleoli by l.)
The nucleoli are very different in respect to their form, size, number and arrange ment. Since these differences are very great even in different specimens of a single species (as, e.g., in the common cosmopolitan Aulacantha scolymantha, Aulosphaera trigonopa, Coelodendrum ramosissimum, &c.), it is probable that they represent different stages of development and multiplication, and that the smallest fragments of the nucleoli, or the final results of their repeated division, become the nuclei of the flagellate spores, which are developed in the PHAEODARIA just as in the other Radiolaria. In the majority of nuclei examined, the number of the enclosed nucleoli proved to be very great, fifty to eighty or more, often some hundreds, the greater their number the smaller their size. Their form is usually irregular, roundish, or even amoeboid- probably the result of amoebiform motions (Pl.101, fig. 1). Sometimes the nucleoli were regularly spherical, equidistant, and connected apparently by a delicate network (Pl.101, fig. 2). (Compare Pl.102, 103, 104, 111, 123, and also Taf. x. of Hertwig, loc. cit.).
The calymma, or the extracapsular jelly-veil, is in the PHAEODARIA always well developed and usually much larger than the enclosed central capsule. The entire volume of the calymma may be three to six times as great as that of the capsule in the majority of this legion; but in the large Aulacanthida, Aulosphaerida, Coelodendrida, Coelographida, &c., the volume of the former is twenty to fifty times as great as that of the latter, or even more. The jelly substance is rather firm and consistent, clear, structureless, and becomes more or less intensely stained by carmine. In the larger forms it is often filled by numerous large alveoles, which are usually absent in the smaller forms. These extracapsular alveoles are most strongly developed in the calymma of the Phaeocystina, or the PHAEODARIA with incomplete skeleton, embracing the three families Phaeodinida, Cannorrhaphida and Aulacanthida (Pls. 101-104). Usually the calymma is here very voluminous and entirely filled up by large alveoles, which are either spherical, irregularly roundish, or polyhedral by mutual compression. These alveoles or vacuoles have no peculiar wall, but are only cavities in the homogeneous substance of the jelly, and are filled by a clear aqueous fluid. Between these the network of the anastomosing pseudopodia is expanded. They exhibit, therefore, the same shape, as was first observed in Thalassicolla and in the Polycyttaria (Collozoida, Sphaerozoida and Collosphaerida).
The relation of the calymma to the skeleton is in the PHAEODARIA of the same importance as in the other Radiolaria, and we may also here distinguish a primary and a secondary calymma. The primary calymma is that on the surface of which at a certain period of life (in the "shell-building period ") the fenestrated shell is secreted in the majority of PHAEODARIA. The secondary calymma, however, is formed after this period, and envelops the shell itself as well as its apophyses externally. Usually the entire skeleton seems to be enveloped by the secondary calymma.
The parts of the extracapsular body, which are enclosed in the gelatinous calymma, possess a peculiar importance in the PHAEODARIA; these are firstly the sarcomatrix and the pseudopodia arising from it, and secondly the phaeodium. The sarcomatrix, or the layer of extracapsular sarcode (ectoplasm), which immediately surrounds the central capsule, is very thick and more strongly developed in all PHAEODARIA than in all other Radiolaria. Its extraordinary size has been already mentioned by Hertwig (1879, loc. cit., p. 99). It is in direct connection with the intracapsular sarcode (or the endoplasm) only by the openings of the central capsule, and mainly by the astropyle. Very numerous radial pseudopodia arise everywhere from the sarcomatrix and run to the surface of the calymma, usually forming a rich network in it by means of numerous branches and anastomoses (compare Pl.101-104). On the surface of the calymma the meshes of this network are very numerous, and there arise from its nodal points the terminal pseudopodia, which float freely in the surrounding water. The metamorphoses of this network of sarcode, the perpetual changes in the number and size of its meshes, and the movements of the sarcode streams as well as of the small granules running in it, are always very manifest in the PHAEODARIA and in the big forms of this legion (mainly in the Aulacanthida) they are better observed than in the majority of other Radiolaria. In many PHAEODARIA (and perhaps in all) a part of the pseudopodia seems to have undergone a local differentiation, for special physiological purposes; and Hertwig has described a peculiar conical contractile body, which arises in Caelodendrum between the two parapylae (loc. cit., p. 100, fig. 3). Further examination of these interesting organs in living PHAEODARIA is required. Usually the sarcode, issuing from the openings of the capsule, forms a stronger cylinder, with peculiar movements.
The phaeodium, or the peculiar dark extracapsular pigment-body of the PHAEODARIA, is one of the most important and most characteristic parts of their organisation, and has induced me to derive their name from it; it is not less typical for the whole legion, than the astropyle with its radiate operculum and the proboscis; and both these important parts are always in direct topographical and physiological connection. Whilst I have missed the phaeodium in no Phaeodarium in which the soft body was well preserved, I have not found it in any other Radiolaria; for the similar extra-capsular pigment bodies, which are found in some species of Thalassicolla and some other Spaerellaria, have a composition and signification different from that of the phaeodiuman exclusive peculiarity of the PHAEODARIA.
The typical importance of the phaeodium for all members of this legion is proved by the following five facts :-( 1) its constant presence in all PHAEODARIA; (2) its constant excentric position in the oral half of the calymma; (3) its constant relation to the astropyle, the operculum and the proboscis of which is always surrounded by it; (4) its constant considerable size, its volume being usually greater than that of the central capsule, the aboral half of which is covered by it; (5) its constant colour and morphological as well as chemical composition. These five facts together demonstrate by their absolute constancy the high morphological and physiological importance of the phaeodium for this peculiar subclass of Radiolaria, although its true nature is difficult to make out, and its main function is not yet sufficiently known.
The first remarks that have been made on the phaeodium were published in 1862 in my Monograph, where I noticed the peculiar dark brown extracapsular pigment body and its excentric position covering only one-half of the central capsule, in Aulacantha (p. 263, Taf. ii. figs. 1, 2), in Thalassoplancta (p. 262, Taf. iii. fig. 10), and in Coelodendrum (p. 361, Taf. xxxii. fig. 1). Its general presence and peculiar composition were frrst recognised by Dr. John Murray, who had, during the Challenger Expedition, the first opportunity of examining many big living PHAEODARIA brought up from great depths. He gives in his first Report (1876, bc. cit., p. 536), the following important notice :-" The sarcode of all these deep-sea Rhizopods has many large black-brown pigment-cells. Small bioplasts are scattered through the sarcode. These collect into capsular-like clumps when the animal is at rest, and are quickly coloured by carmine." In 1877 Dr. John Murray sent me the wonderful collection of PHAEODARIA brought home by the Challenger, and I had now the best opportunity for examining the phaeodium in hundreds of well-preserved specimens. Supported by these extensive observations, I gave, in 1879, in a preliminary paper,1 a fuller description of the phaeodium, and of the phaeodella (or the peculiar dark pigment-granules composing it). At the same time R. Hertwig published his observations on the big living TRIPYLEA examined by him at Messina, and pointed out particularly that the constant presence, composition, and arrangement of this excentric extracapsular pigment-body was most characteristic, and sufficient in itself to distinguish this group from all the other Radiolaria (1879, loc. cit., p. 99).
The most striking peculiarity of the phaeodium, and the most important on account of its absolute constancy, is its excentric position, covering only the oral hemi sphere of the central capsule, and wholly or partly wanting on the aboral hemisphere. This constant topographical relation to the capsule never fails, and may be always observed at the first glance, when the body is seen from the side (the main axis of the capsule being perpendicular to the axis of the eye of the observer), (compare Pl. 101, figs. 6, 10; Pl.102, fig. 1; Pl.103, fig. 1, &c.). The phaeodium envelops, therefore, the oral half of the capsule completely, and especially the astropyle in its centre; hence, the radiate operculum and the proboscis arising from the latter cannot be seen usually before the former is removed. The general form of the entire phaeodium, in consequence of this excentric position, is concavo-convex (or crescentic in longitudinal section), its concave face embracing the oral or anterior face of the capsule, and its convex face being turned to the surface of the calymma.
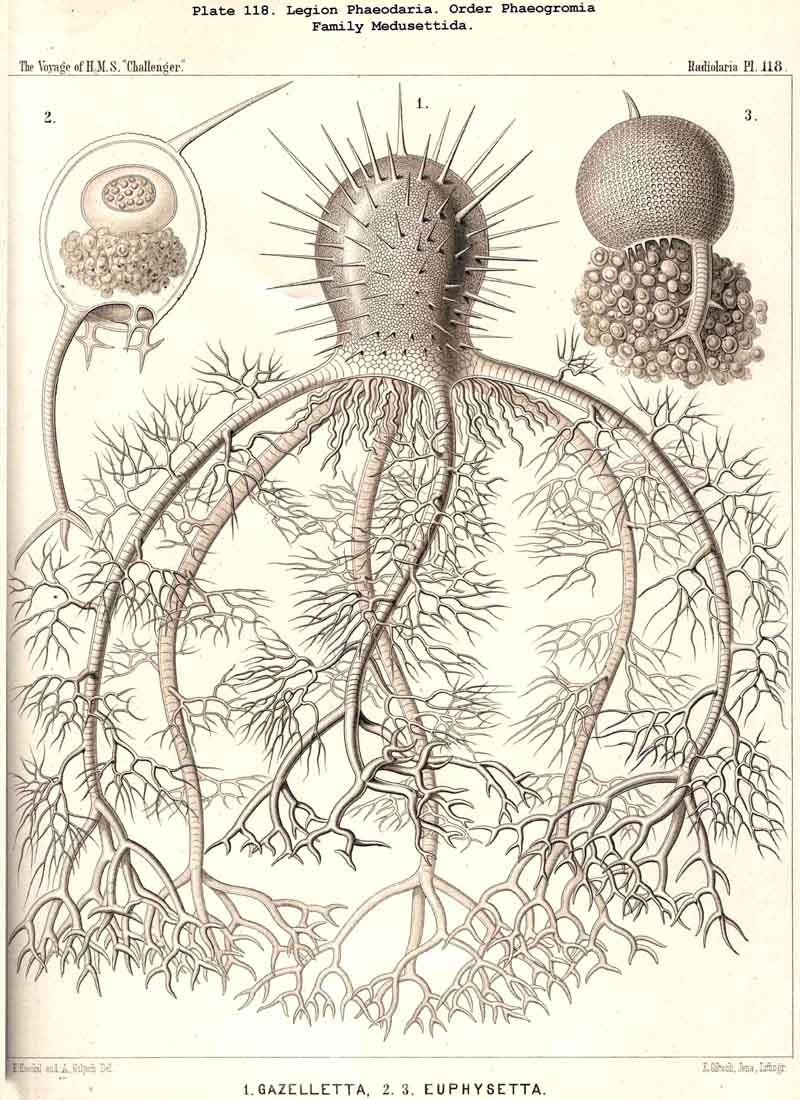
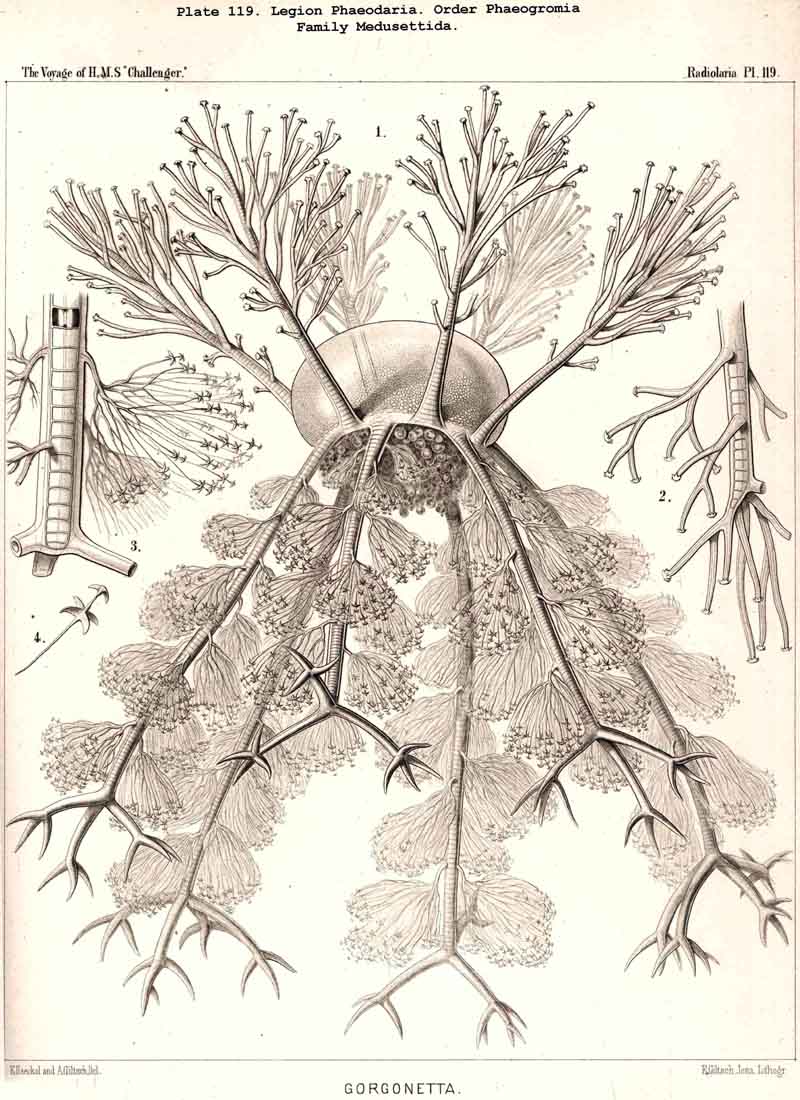
The topographical relation of the phaeodium to the surrounding shell is also very characteristic in the suborder Phaeogromia, or in those PHAEODARIA which possess a peculiar shell-mouth placed on the oral pole of its main axis. Here the capsule is always placed in the aboral half of the shell-cavity, the phaeodium in its oral half, separating the astropyle from the mouth of the shell, both of which lie in the main axis; as in the Challengerida (Pl.99), Tuscarorida (Pl.100), Castanellida (Pl. 113), Circoporida (Pl.115), and Medusettida (Pl.118, 119, 120). In this suborder (the Phaeogromia), the volume of the phaeodium may be, on an average, about as great as that of the central capsule, whilst in the majority of other, PHAEODARIA it is much greater than the latter.
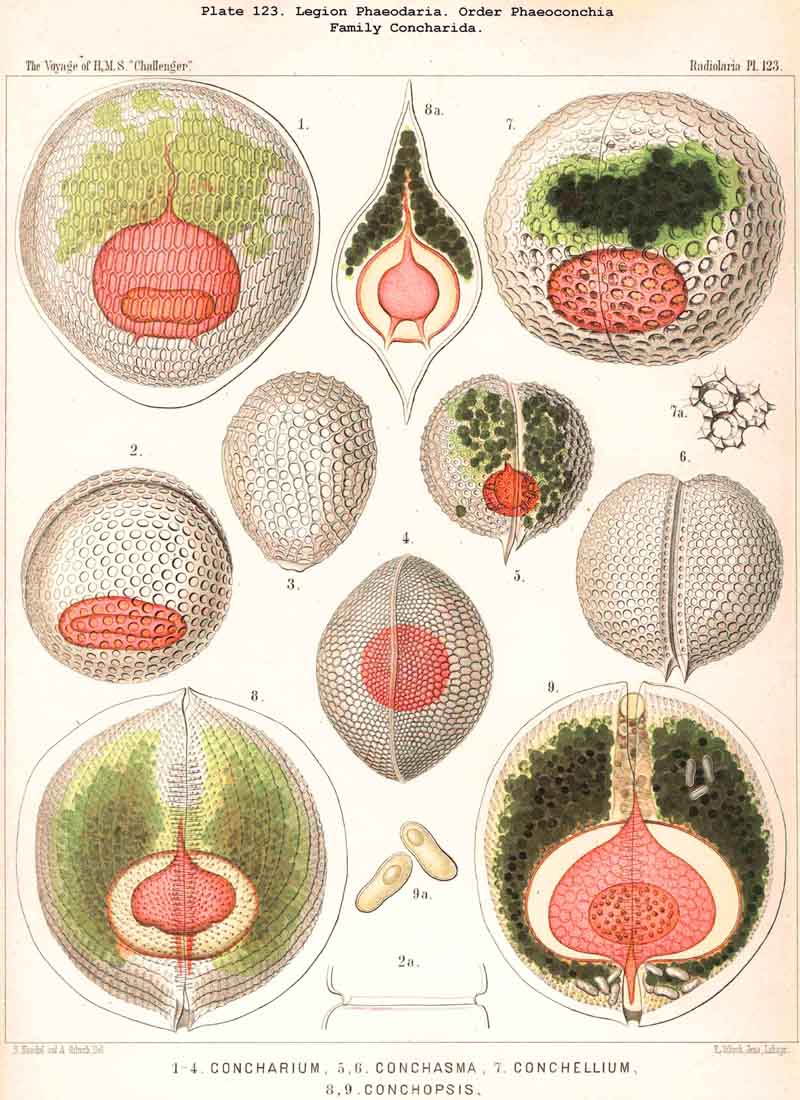
A similar topographical relation between the phaeodium and the enclosing shell, as in the Phaeogromia, also exists in the suborder Phaeoconchia, or in those PHAEODARIA, which are distinguished from all the others by the possession of a bivalved shell (Pl. 121 - 128). The two valves of this curious shell, which resembles that of the Brachiopoda, are dorsal and ventral, and the tripylean central capsule is always so placed between them that its two lateral parapylae (right and left) lie in the frontal plane of the shell, where a large frontal fissure opens between the opposed margins of the two hemispherical or cap-shaped valves. The phaeodium is also placed here on the oral half of the capsule and surrounds its astropyle; but it exhibits some differences in the three families of Phaeoconchia.
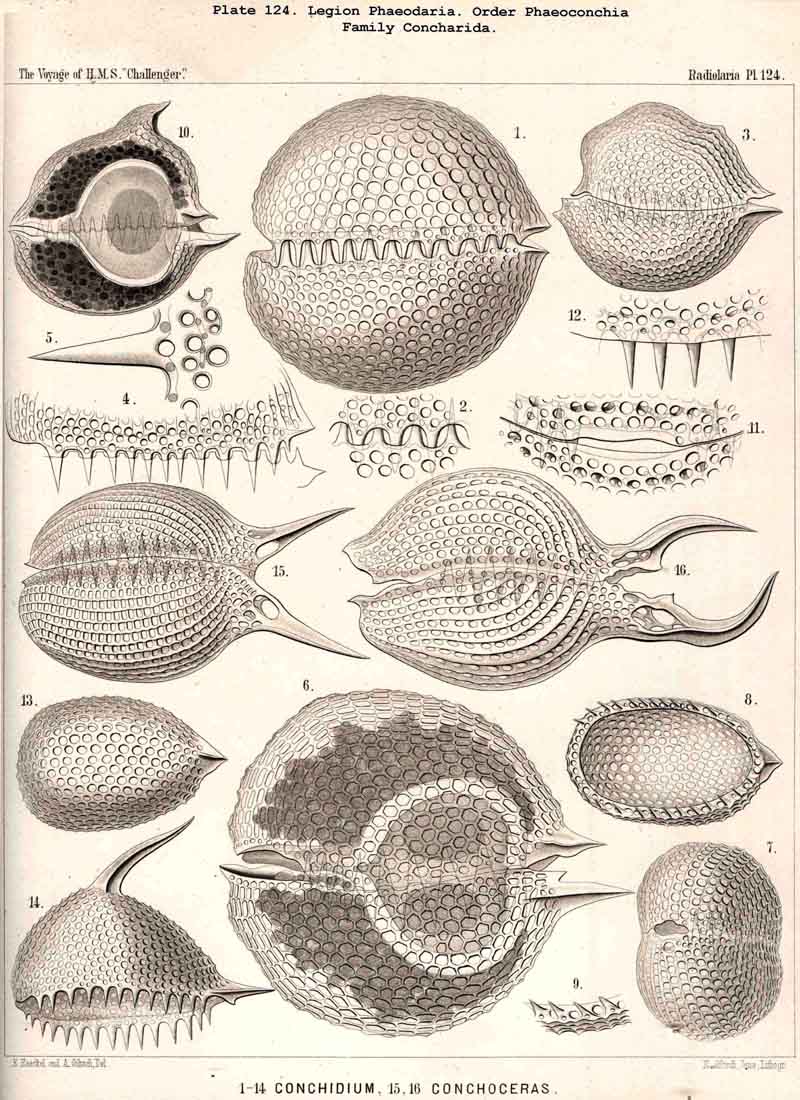
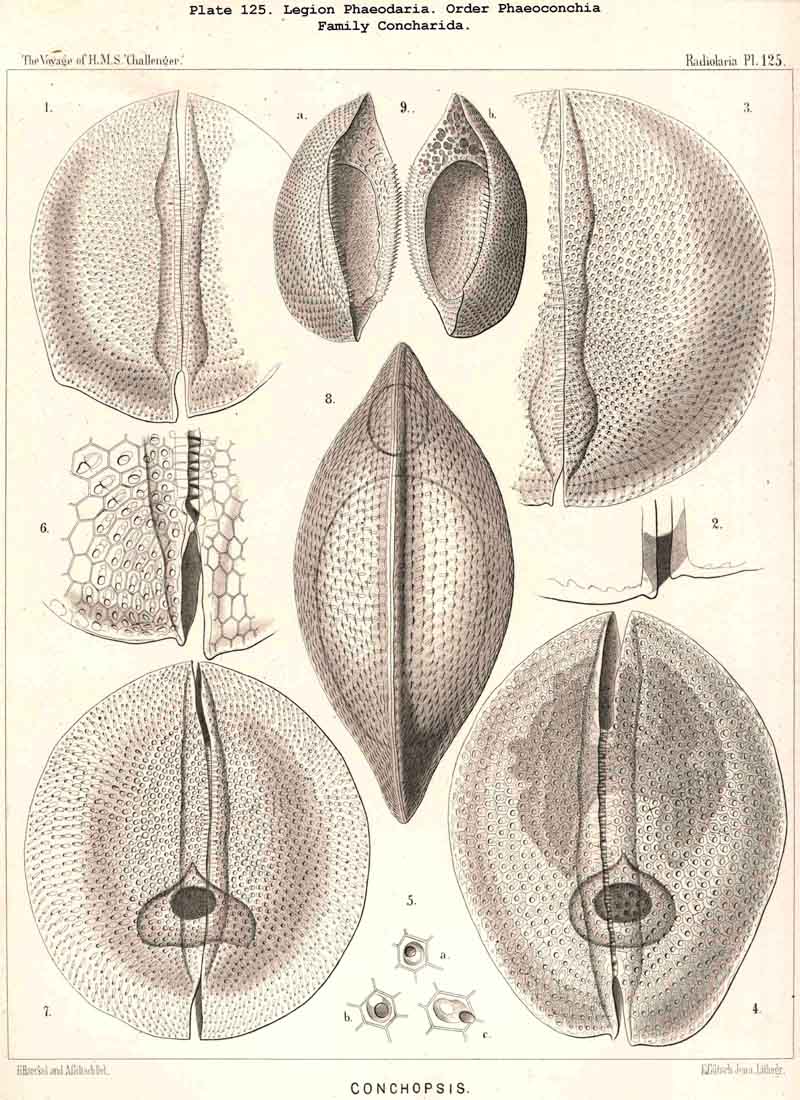
The Concharida, the bivalved shell of which is simple, and without tubular apophyses (Pl.123, 124, 125), possess a relatively small central capsule, which usually fills up only the third or fourth part of the shell-cavity. This is the aboral or posterior part, on the apex of which both valves are united by a ligament in some Concharida (Pl.123, figs. 8, 9). The oral or anterior part of the shell-cavity (usually two-thirds or more) is filled up by the phaeodium, and this is usually bifid, being divided by a frontal constriction into two wings or lobes; the dorsal wing is hidden in the upper valve of the shell, the ventral wing in the lower valve; both wings are usually united only by a small central bridge, and this bridge of the phaeodium is pierced in its centre by the proboscis of the astropyle (Pl. 124, figs. 6, 10; Pl. 123, figs. 8, 9).
The Coelodendrida have a different shape (Pl. 121). Their bivalved shell is relatively small and tiny, and bears on the two poles of the sagittal axis two conical apophyses or galeae, from each of which three or four very large, dichotomously branched tubes arise. The central capsule fills up the cavity of the bivalved shell almost entirely, and the voluminous dark phaeodium envelops both to such an extent that the shell and the enclosed capsule are often hidden in it completely. Therefore I arrived in my first description of Coelodendrum (1862, bc. cit.) at the erroneous conclusion that the capsule lies outside, not inside the shell. The first accurate figure and description of its structure was given in 1879, by Hertwig (loc. cit., p. 99, Taf. x. fig. 3). The central capsule (v) is here separated from the bivalved shell (m) only by a very small distance, and the oral part of both is hidden in the phaeodium. I find, however, in the majority of the numerous preparations of the Challenger collection, the volume of the phaeodium much greater, and it often envelops the entire shell.
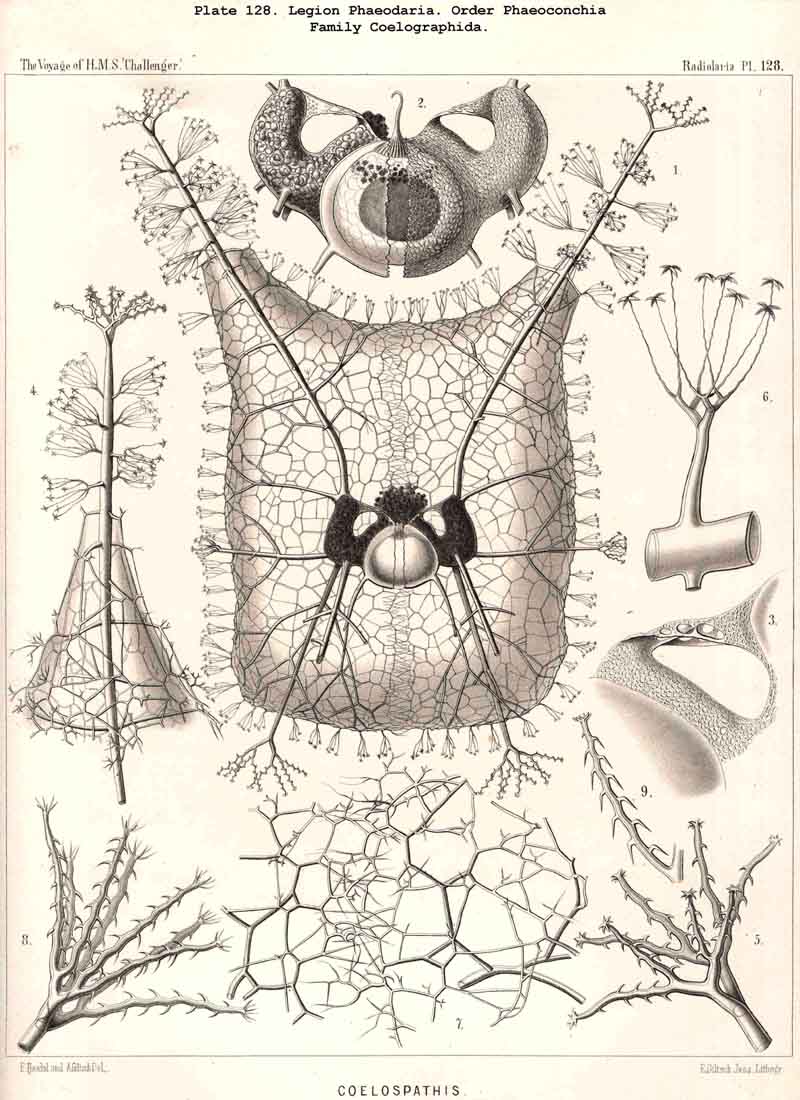
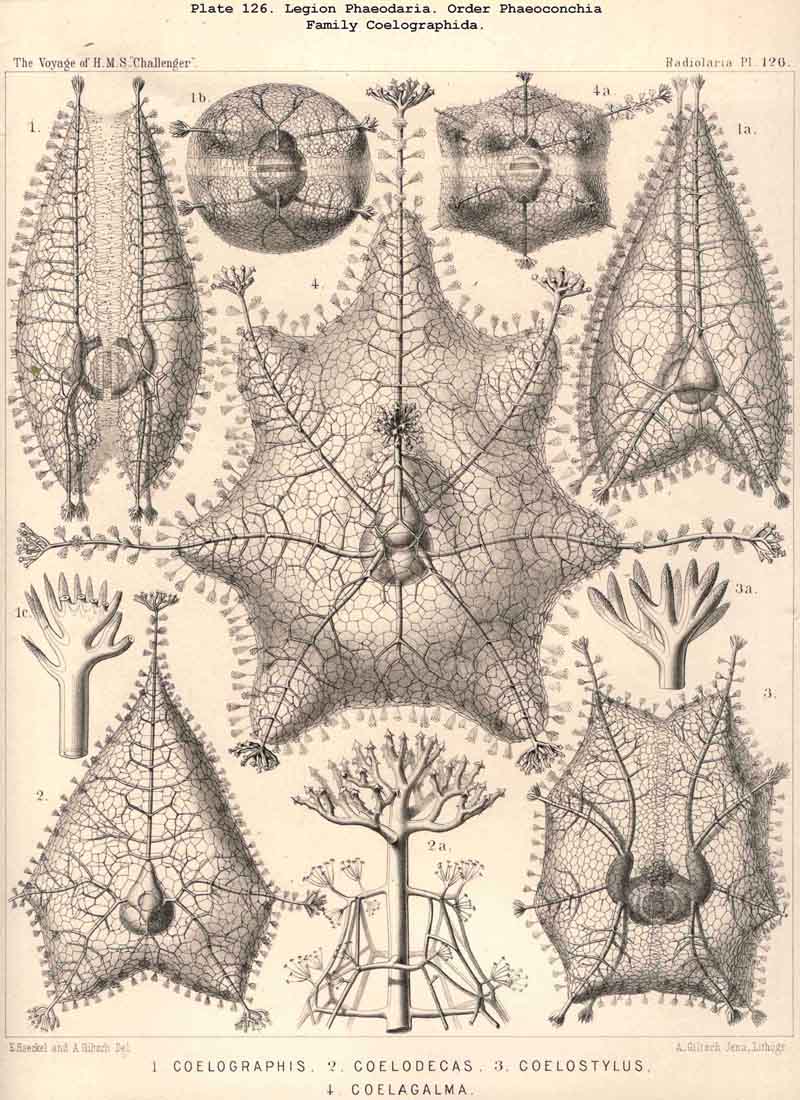
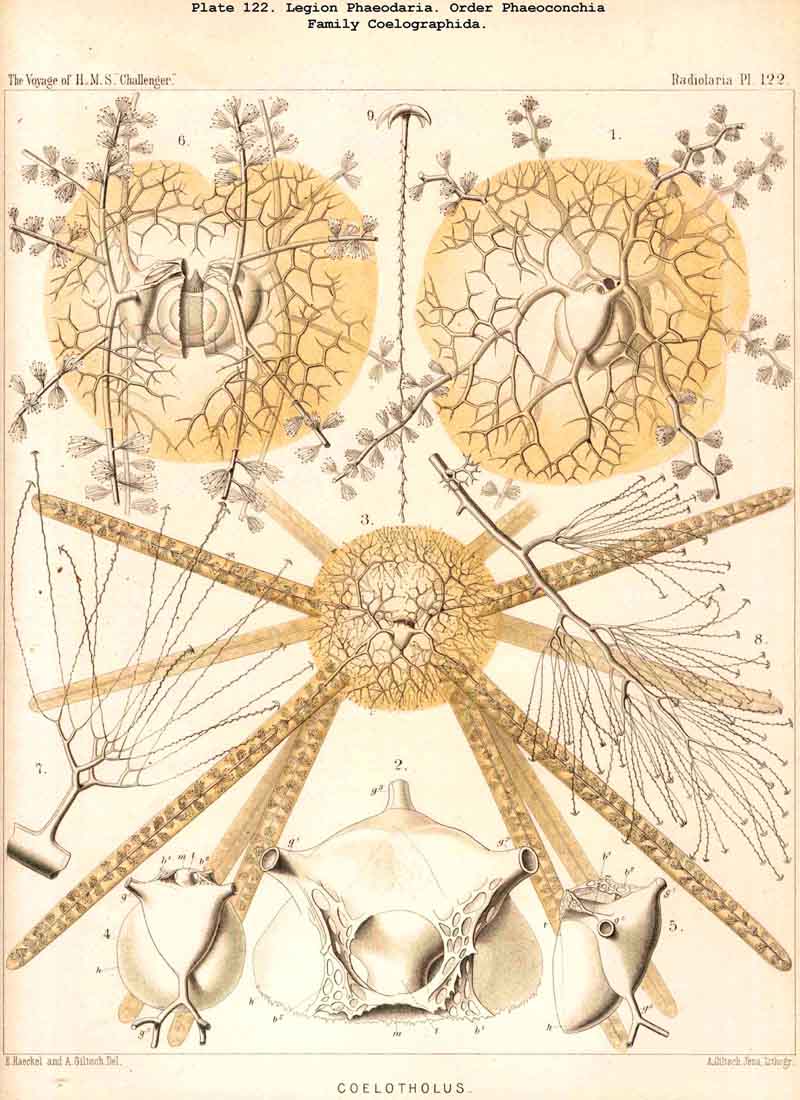
The Coelographida, finally, have a phaeodium of the most remarkable shape, since in their bivalved shell a peculiar reserve store or magazine of phaeodella, which we call the "phaeocapsa" is developed for it (Pl.126, 127, 128, g.t.m.). The bivalved shell has in these most perfect PHAEODARIA a structure similar to that in the Coelodendrida; but they differ from the latter in the stronger development, and greater differentiation of the two apical galeae, and the large hollow tubes arising from them. These two helmet-shaped cupolae, the galeae (g), which arise from the two valves on the poles of the sagittal axis, are in the Coelographida usually larger than the valves themselves, and are not closed, as in the Coelodendrida, but open by a tubular apophysis at their base, the nasal tube or rhinocanna (t). The apex of the galea is connected with the open mouth of the rhinocanna by a single or double frenulum (b). The two nasal tubes or rhinocannae (a dorsal and a ventral) lie in the sagittal plane of the body (and run from the base of each galea along the anterior convexity of the valve to its oral margin. Here is placed the proboscis of the astropyle, between the two opposed mouths of the rhinocannae (Pl. 128, fig. 2). The phaeodium is usually hidden entirely in the two phaeocapsae, which are composed of the two galeae (g), and the two rhinocannae arising from them (Pl.127, figs. 4-9). A part of the phaeodella is usually thrown out by the mouth of the latter (m).
The characteristic colour of the phaeodium exhibits numerous different tints between green, brown, and black. It seems to be in the majority blackish-brown or greenish- brown, very often olive, more rarely almost quite green or red-brown. Usually the colour is so dark, intense and opaque, that the parts enclosed by the phaeodium, mainly the oral hemisphere of the central capsule and the astropyle, are completely hidden in it. The chemical composition of the phaeodium demands further accurate researches; unfortunately I have not been able to make out its true nature, since numerous different experiments furnished no certain general results.
The phaeodella, or the pigment-corpuscles, which compose the phaeodium, aggregated in hundreds, and in the bigger species in thousands, are usually spherical, some times somewhat ellipsoidal, at other times spheroidal or lenticular; but usually numerous smaller, irregular, roundish particles are intermingled between the larger and more regular corpuscles, and often the main mass forms a very fine black powder. The size of the phaeodella is very variable, not only in the different species, but also in one and the same individual. The larger phaeodella have a diameter of 0.01 to 0.02, the smaller of 0.004 to 0.008 mm.; but there also occur very big forms of 0.04 to 0.05 mm., or even more, and very small ones of 0.001 mm. or less. Usually the phaeodium appears as an aggregate of numerous larger and smaller phaeodella, which are very different in size as well as in the intensity and tint of their colour, and are irregularly crowded in a black, powder-like substance.
The morphological nature of the phaeodella is also difficult to make out. I have already pointed out in my first description of Aulacantha, Thalassoplancta and Coelodendrum (1862, loc. cit.), that a great part of these pigment-corpuscles are true cells, composed of a nucleus and protoplasm, which contains granules of pigment, and is enveloped by a membrane. Dr. John Murray, who had during the Challenger voyage the opportunity of examining numerous different living PHAEODARIA, and staining them by carmine, also asserts that a great part of those dark corpuscles are "large black-brown pigment-cells" (1876, loc. cit., p. 536). Numerous preparations of the Challenger collection, well preserved in glycerine, and stained by carmine, contain PHAEODARIA belonging to different families, the phaeodium of which contains numerous such "pigment-cells," with a dark red nucleus, and so similar are these cells, that every histologist should recognise them. But in strange contrast to this is the fact, that in numerous other mountings, prepared in the same manner, not a single cell of this kind is found in the phaeodium, and that the latter is composed only of irregular pigment-granules. In many PHAEODARIA belonging to different families I, like Hertwig, could not find a single true nucleated cell in the phaeodium.
A great part of the PHAEODARIA, and usually the bigger forms of Aulacanthida, Cannorrhaphida, Coelodendrida, Coelographida, &c., exhibit a peculiar structure of the larger phaeodella, viz., a fine parallel striation (Pl. 101, figs. 3, 6; Pl.102, fig. 1; Pl.103, fig. 1; Pl.104, figs. 1-3, &c.). In each phaeodellum may be counted about ten to twenty such fine parallel stripes (more in the greater, less in the smaller forms); and in the ellipsoidal phaeodella the stripes are either transverse rings, perpendicular to their main axis, or ascending obliquely; they often resemble the convoluted spiral filament of a thread-cell or nettle-cell of an Acaleph. Some times these parallel transverse stripes are very striking. Another structure is seen in larger phaeodella, namely an aggregate or cluster of smaller globules, often of equal size, resembling a small morula. All these minute structures of the phaeodella as well as their changes in the living PHAEODARIA, require a far more extensive examina tion (by means of strong lenses and different chemical reagents), than I could, unfortunately, devote to them.
The physiological signification of the phaeodella, therefore, is at present not yet known; but the general facts quoted above, their constant presence, position, volume, and composition, make it probable that their physiological value in the PHAEODARIAN organism is very great. The following hypotheses may be taken provisionally into consideration :- A. The phaeodella are peculiar symbiontes, or unicellular algae, comparable to the xanthellae or zooxanthellae of the other Radiolaria. This hypothesis is probably correct for those phaeodella which are true nucleated cells ; and the more so, as the majority of PHAEODARIA do not exhibit those common yellow xanthellae, which are usually found in the SPUMELLARIA and NASSELLARIA. It is even possible that the latter are absent in all PHAEODARIA. B. The phaeodella are dark pigment-bodies, which absorb light and heat in a manner similar to the simple "pigment-eyes" of many lower animals, and may therefore be optical sense-organs of the PHAEODARIA. This hypothesis may be supported by a comparison with the large-eyed unicellular Protist, Erythropsis agilis,described by R. Hertwig. C. The phaeodella are organs of nutrition of the PHAEODARIA and active in their metastasis (" Stoffwechsel "). Regarding them from this point of view, we may suppose that the phaeodella are secreted products which serve for digestion, acting like the bile or the saliva of higher animals. Perhaps they too act like the venomous matter produced in the thread-capsules of the Acalephae. The suggestion that they are mere excretions, or half-digested matters, as Hertwig supposes (" halb assimilirte Nahrungs-bestandtheile," 1879, loc. cit., p. 99) seems less probable. The most important fact illustrating their high signification for the processes of nutrition, digestion and for effecting changes on matter, seems to be the close relation of the phaeodium to the astropyle; the radiate operculum of the latter, and the proboscis arising from it, being constantly covered and completely hidden by the central main mass of the phaeodium.
The skeleton of the PHAEODARIA is always extracapsular, and exhibits in the majority of this legion such a characteristic shape, form, and structure, that these organisms may be easily recognised by it, even apart from the central capsule and the phaeodium. In a few cases, however, the skeleton is so similar to that of some NASSELLARIA and SPUMELLARIA, that it may be accidentally confounded with it. In general the skeleton of the PHAEODARIA is much larger, and much more highly developed, than that of most other Radiolaria, and exhibits the most wonderful appearances, and the most marvellous complications, which are found in the whole world of Protists, or of unicellular organisms. The varied composition and differentiation of the skeleton alone distinguishes the numerous families, genera, and species of PHAEODARIA described in the sequel; all the fifteen families, however, agree so comPletely in the structure of the central capsule and the phaeodium described, that we may derive them all phylogenetically from a small skeletonless family, the Phaeodinida.
The chemical composition of the skeleton seems to be, in the majority of PHAEODARIA, somewhat different from that of the other Radiolaria. In a few groups only, especially in the Cannobelida (Dictyocha, Mesocena, &c.), and in a part of the Castanellida and Concharida, the substance of the skeleton seems to be of pure silica, as in the NASSELLARIA and SPUMELLARIA; these flinty skeletons, therefore, may be also found fossil. In the majority of PHAEODARIA, however, the skeleton does not consist of pure silica, but of an organic silicate; it becomes more or less intensely stained by carmine, and browned or blacked by fire; in many cases it even becomes comPletely burned and destroyed by the prolonged action of heat. This circumstance explains why PHAEODARIA in general are rare in deep-sea deposits, as in the common Radiolarian ooze of the Pacific, and why they are generally absent in fossil deposits. Even the pure Radiolarian rocks of the Barbados, &c., contain only a few PHAEODARIA, mainly Dictyochida.
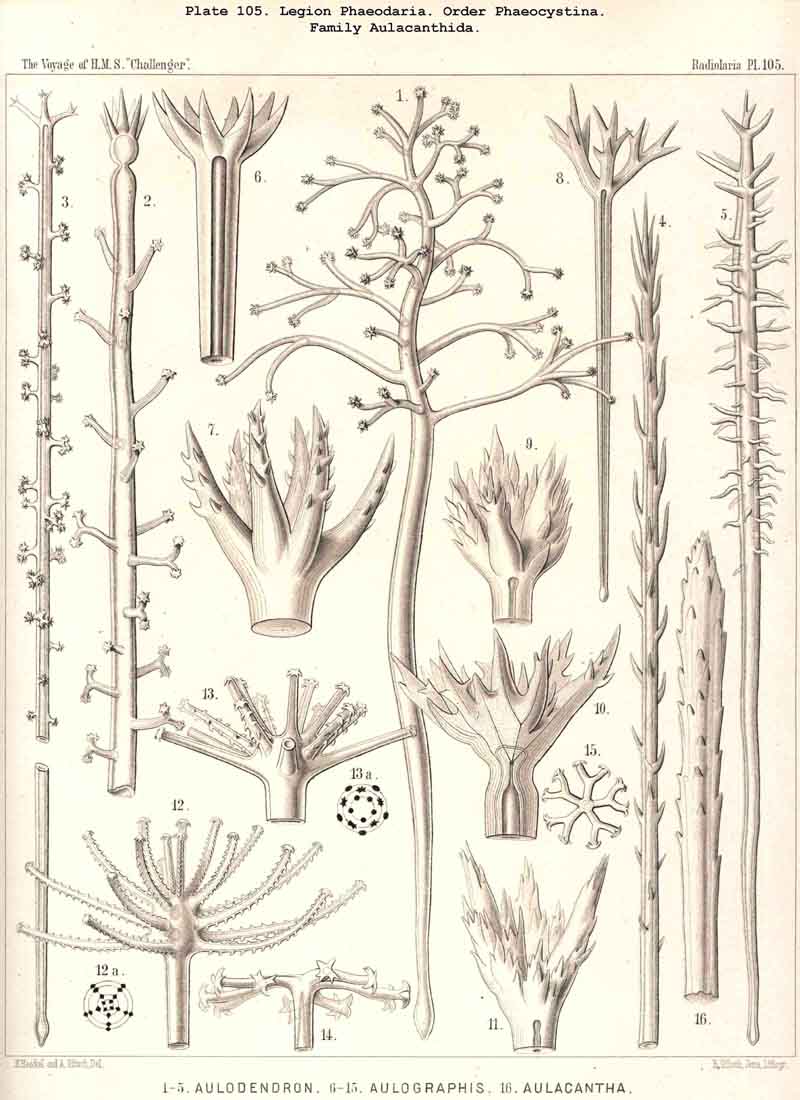
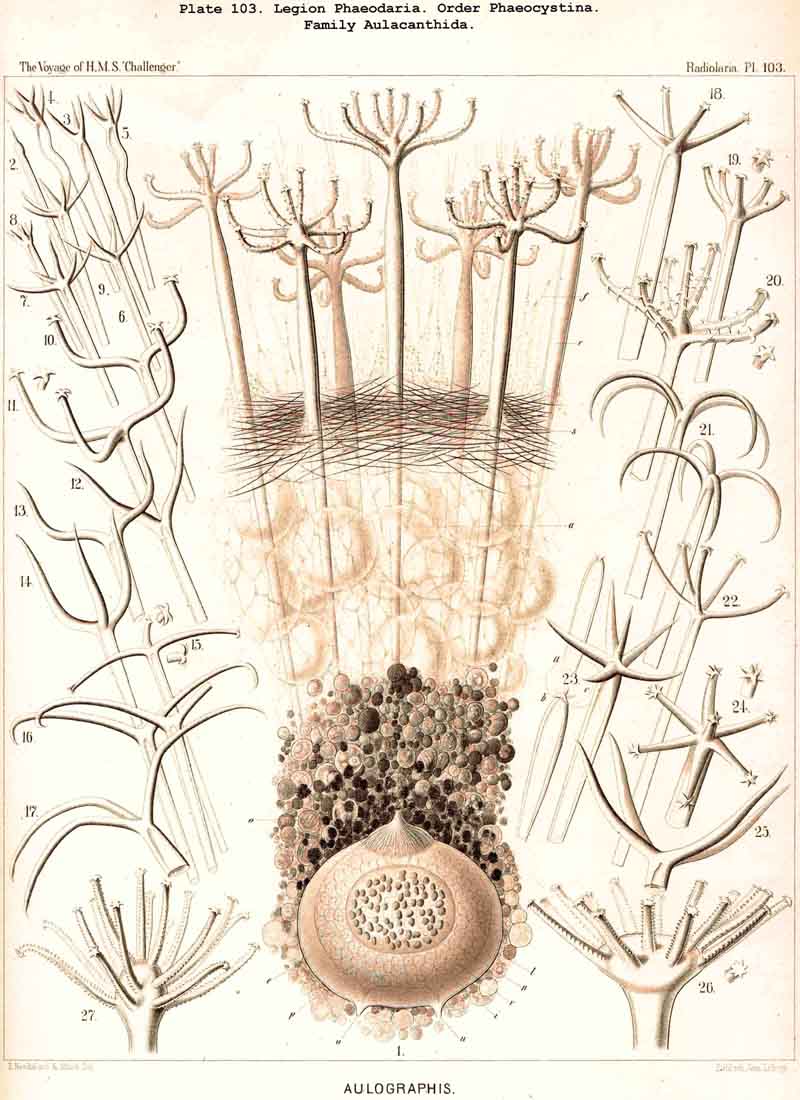
According to the different forms of the skeleton, we may divide the legion or sub class of PHAEODARIA into two sublegions, four orders, and fifteen families. Firstly, we may distinguish as two groups the Phaeocystina, without a lattice-shell, and the Phaeocoscina, with a lattice-shell (compare above, p. 5). The Phaeocystina comprise three different families, viz., (1) Phaeodinida, without any skeleton (Pl.101, figs. 1, 2); (2) Cannorrhaphida, with an incomplete skeleton, composed of numerous separate, not radial pieces, which are scattered around the capsule in the calymma (Pl.101, figs. 3-14; Pl.114, figs. 7-13), and (3) Aulacanthida (Pl.102-105), with an incomplete skeleton, composed of numerous hollow radial tubes, which pierce the calymma and come in contact by their proximal ends with the surface of the central capsule.
The Phaeocoscina, or the PHAEODARIA with a lattice-shell (embracing the great majority of the whole legion) exhibit three principal differences in the shape of their shell, and from these we distinguish the three following orders; (A) Phaeosphaeria, with a spherical, not bivalved shell (rarely of an ellipsoidal or lenticular, or another modified form), without a shell-mouth or a peculiar constant large opening on the lattice-shell; (B) Phaeogromia, with an ovate or polyhedral, not bivalved shell (often also of a subspherical, ellipsoidal, or another modified form), constantly provided with a shell-mouth or a peculiar large opening on one pole of the main axis of the lattice-shell; (C) Phaeoconchia, with a bivalved shell, composed of two completely separated, hemispherical, cap-shaped or boat-shaped valves (a dorsal and a ventral), comparable to that of the Brachiopoda.
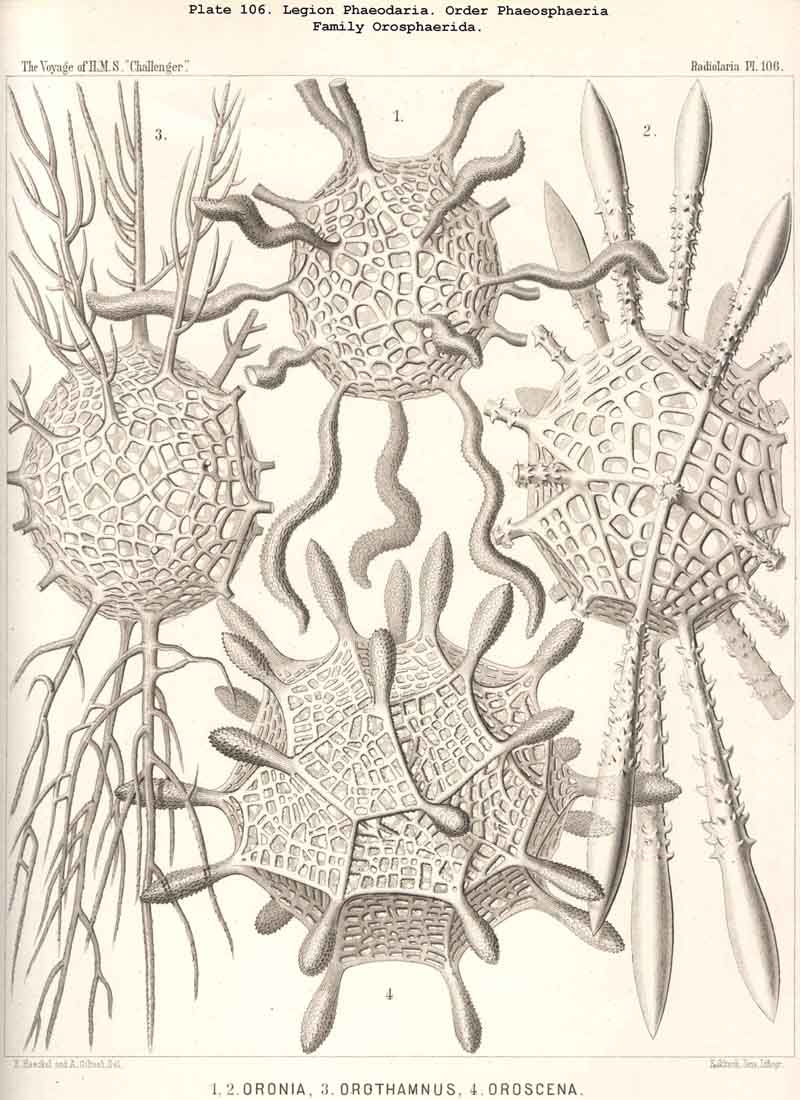
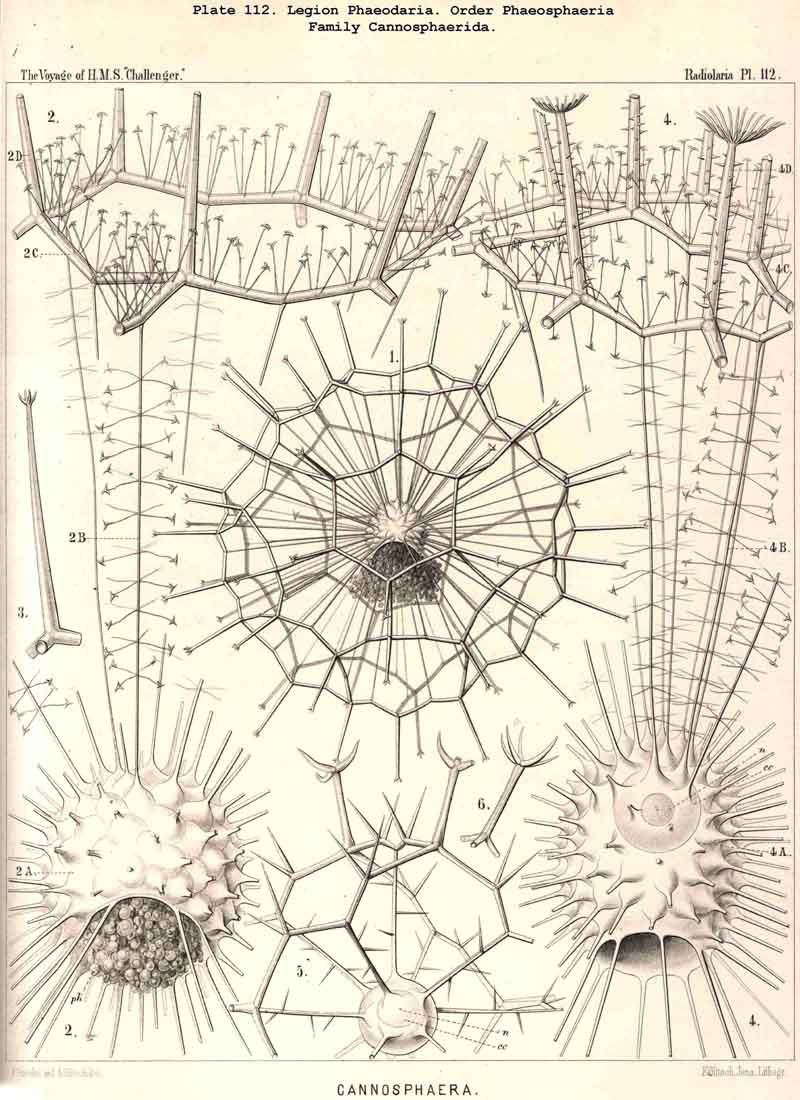
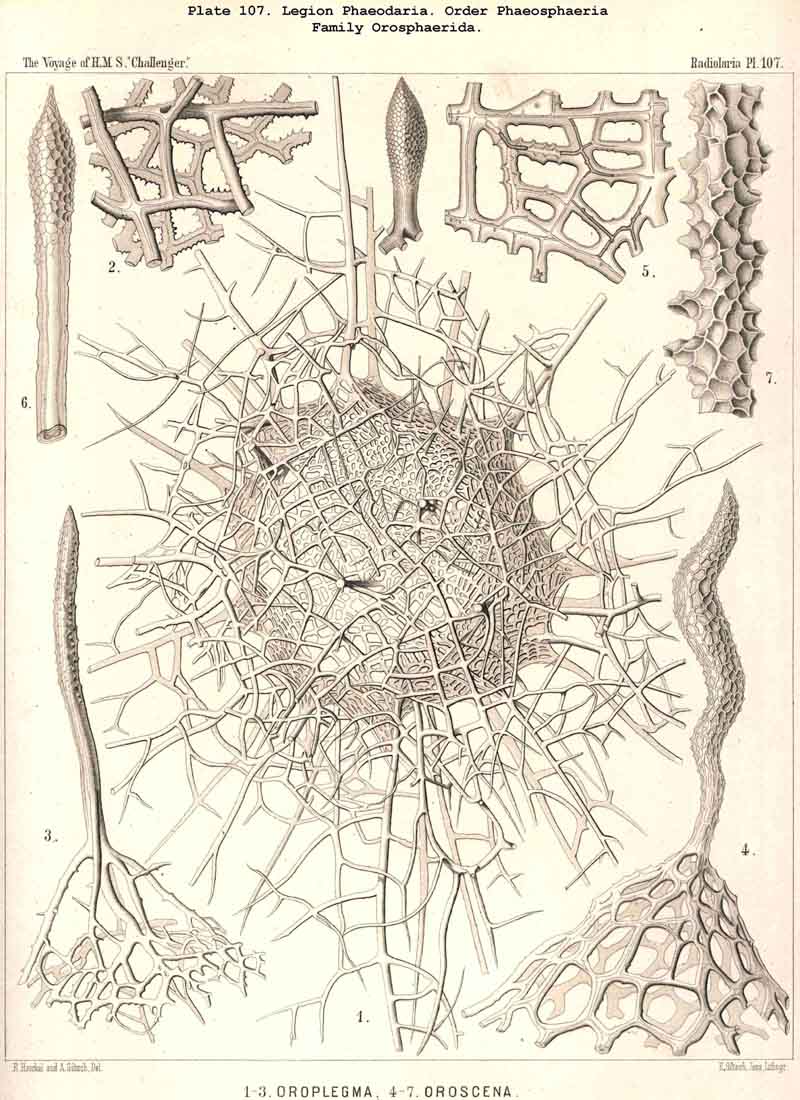
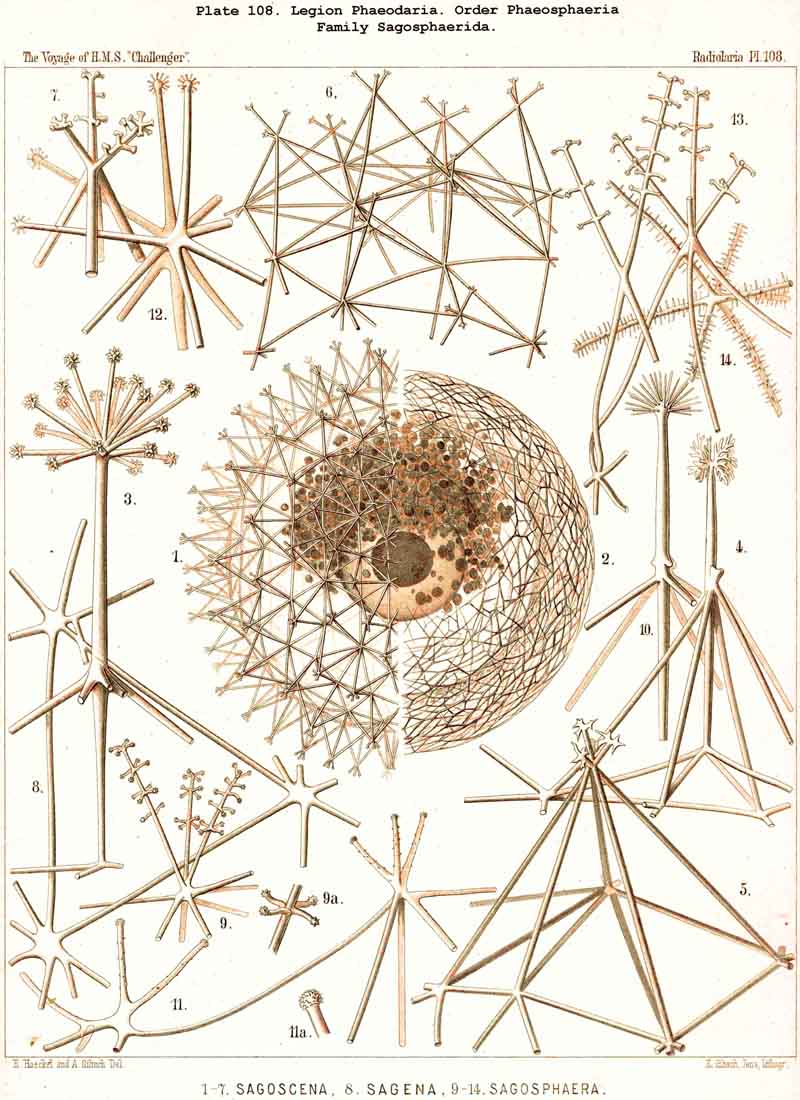
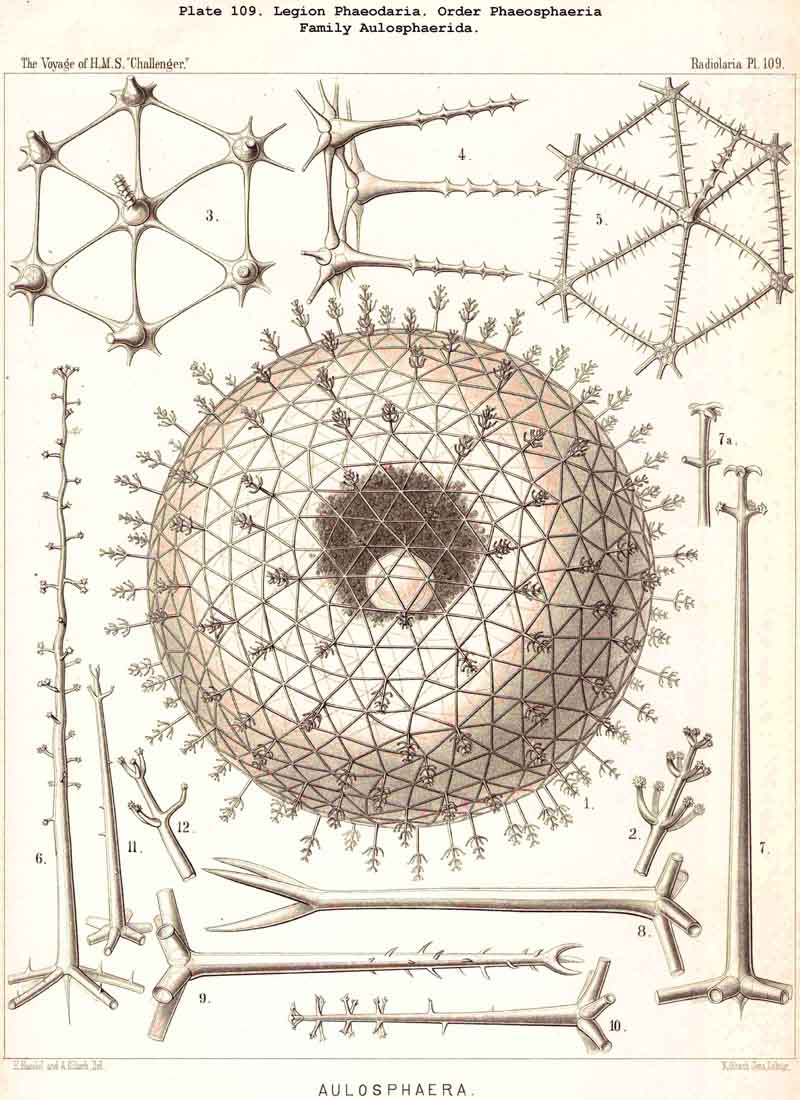
The Phaeosphaeria, or those PHAEODARIA the big shell of which is usually spherical, never bivalved and never provided with a peculiar shell-mouth, comprise a great number of common and large-sized PHAEODARIA, which may be arranged into four different families, according to the different structure of the shell-( 1) Orosphaerida (Pl.106, 107), spherical shell extremely big and robust, composed of a single piece of coarse lattice- work, the thick bars of which are stratified and contain partly a fine axial-canal, meshes of the network usually irregularly polygonal, no astral septa in the nodal points; (2) Sagosphaerida (Pl.108), spherical shell large-sized, but extremely delicate and fragile, composed of a single piece of arachnoidal lattice-work, the thin bars of which are simplesolid threads, without axial-canal, meshes of the network always large and triangular, no astral septa in the nodal points; (3) Aulosphaerida (Pl.109 - 111), spherical shell large-sized, but very fragile, composed of numerous hollow cylindrical tubes, which are connected (and at the same time separated) by peculiar astral septa in the nodal points, meshes either triangular or polygonal; (4) Cannosphaerida (Pl.112), spherical shell double, composed of two concentric shells which are connected by thin hollow radial tubes, the inner shell simPle, solid or fenestrated, with a shell-mouth on the basal pole, the outer shell composed of hollow cylindrical tubes which are connected by astral septa in the nodal points. The structure of this outer shell is the same as in the Aulosphaerida, while the basal mouth of the inner shell brings this family in closer relationship to the Phaeogromia.
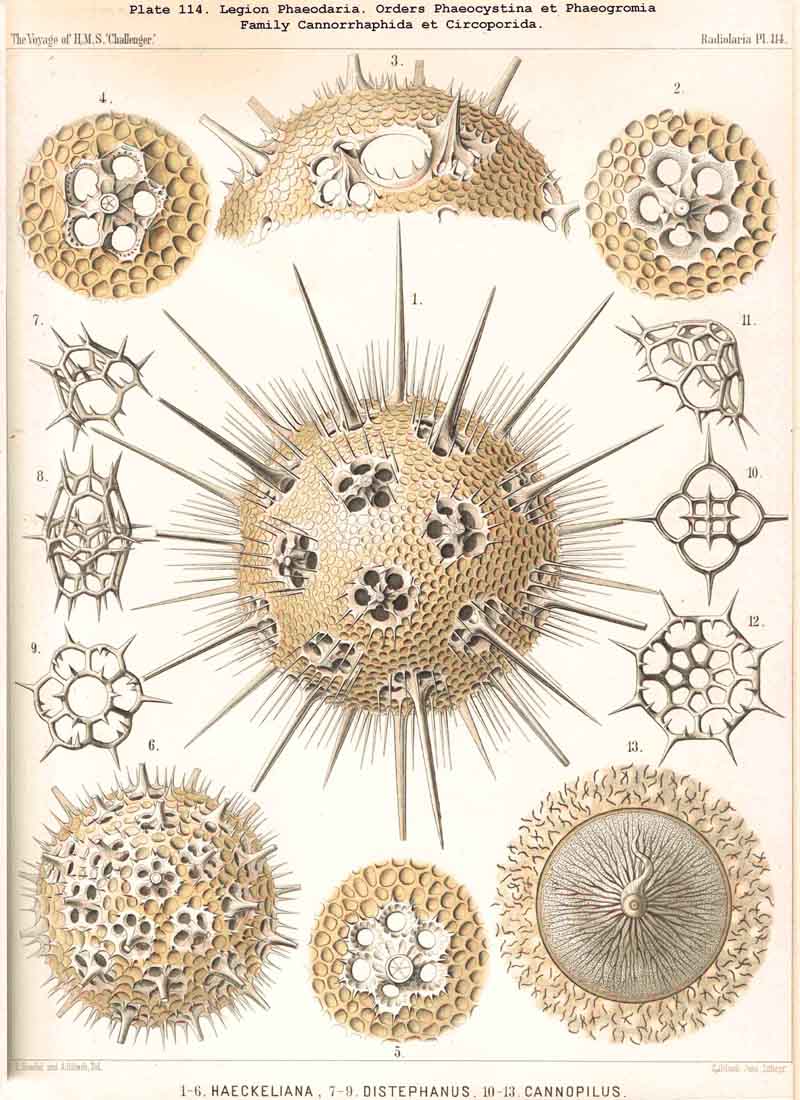
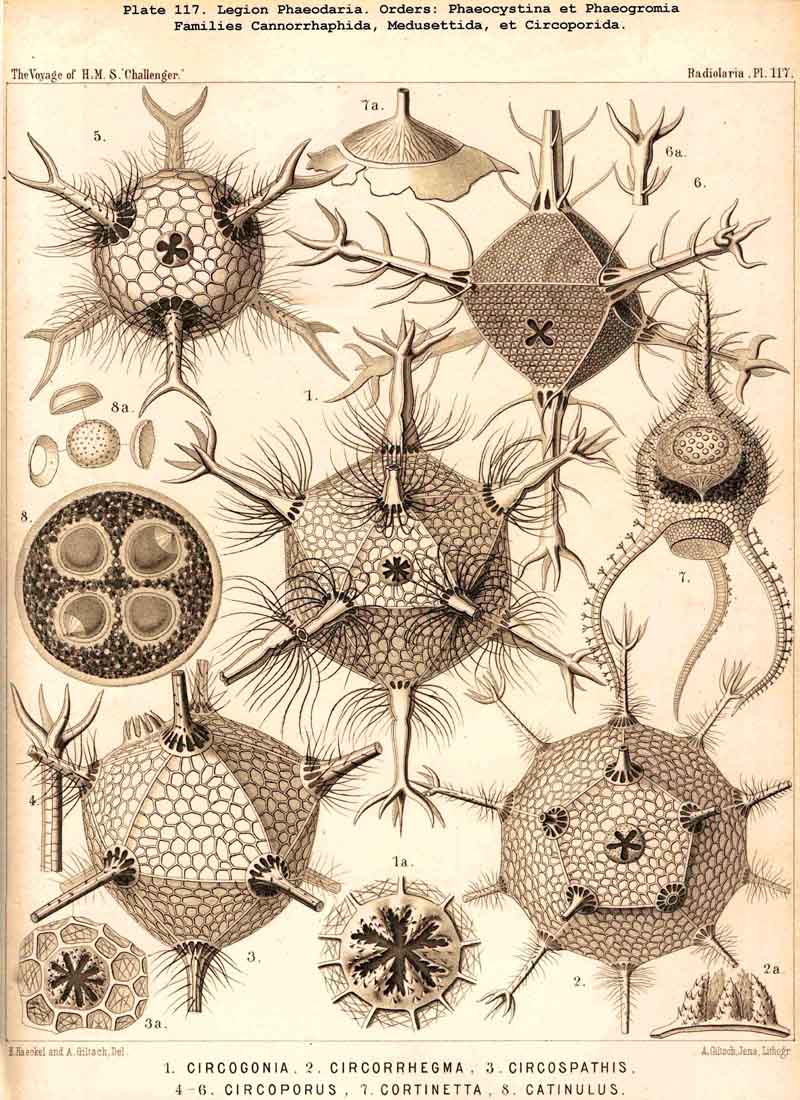
The Phaeogromia, or those PHAEODARIA the shell of which is not bivalved, but provided with a peculiar constant mouth on the oral pole of the main axis, are in general similar to the NASSELLARIA (Monocyrtida), and may be divided into five different families, viz., (1) Challengerida (Pl. 99), shell ovate or subspherical, also often triangular or lenticular, distinguished by a peculiar diatomaceous structure, an exceedingly fine tracery of regular hexagonal, very delicate network; (2) Medusettida (Pl. 118 -120), shell ovate, campanulate or cap-shaped, distinguished by a peculiar alveolar structure, with a corona of peculiar hollow, large, articulated feet around the mouth; (3) Castanellida (Pl.113), shell spherical or subspherical, of ordinary simple lattice-work, usually with a corona of simplesolid teeth around the mouth; (4) Circoporida (Pl.114 -117), shell spherical or polyhedral, with panelled or dimpled surface, distinguished by a peculiar porcellanous structure (numerous thin needles being embedded in a punctulate cement-substance), with hollow radial spines and with a corona of simplesolid teeth around the mouth; (5) Tuscarorida (Pl.100), shell ovate or subspherical, with smooth surface, of the same peculiar porcellanous structure as the Circoporida, but with hollow, very long, tubular teeth around the mouth.
The Phaeoconchia are the peculiar and most interesting "PHAEODARIA bivalva," differing from all other PHAEODARIA, and from all known Radiolaria in general, in the possession of a bivalved lattice-shell, composed of a dorsal and a ventral valve. They may be divided into three families : (1) Concharida (Pl.123, 124, 125), shell with two thick and firm, regularly latticed valves, which bear no hollow tubes and no cupola or galea on their apex or sagittal pole; (2) Coelodendrida (Pl.121), shell with two thin and fragile, scarcely latticed valves, which bear a conical cupola or a helmet-shaped galea on their apex, and hollow branched tubes arising from it (without rhinocanna and frenula); (3) Coelographida (Pl.122, 126, 127, 128), shell with two thin and fragile, scarcely latticed valves, similar to those of the Coelodendrida, but differing from them in the development of a peculiar rhinocanna or nasal tube upon each valve; this tube is connected by an odd or paired frenulum with the apex of the galea, and both together contain the phaeodium.
The phylogenetic affinity of the fifteen families enumerated, and the morphological relationship based upon it, form a very difficult problem. The whole legion of PHAEODARIA is probably monophyletic, in as much as all the families may be derived from a single ancestral group, the skeletonless Phaeodinida (Phaeodina and Phaeocolla); but at the same time polyphyletic, in as much as probably many families have been derived, indepen dently one from another, from different branches of Phaeodinida; or in other words, the characteristic malacoma of the PHAEODARIA (the cannopylean central capsule and the calymma with the phaeodium) may be a monophyletic product, inherited from a single ancestral form; the manifold skeleton, however, is certainly a polyphyletic product, originating from different skeletonless Phaeodinida.
Among the independent families of PHAEODARIA, derived directly from skeletonless Phaeodinida by production of a peculiar skeleton, may be the following : Cannorrhaphida (Pl.101, probably polyphyletic) Aulacanthida (Pl.102-105), Castanellida (Pl.113), Challengerida (Pl.99), Concharida (Pl.123, 124, 125), Circoporida (Pl.114 -117) and Tuscarorida (Pl.100). The four families of Phaeosphaeria (the Orosphaerida, Sagosphaerida, Aulosphaerida and Cannosphaerida (Pl.106-112), may be derived perhaps from the Castanellida; and the Medusettida (Pl.118 -120), have been perhaps derived from the Challengerida. The complicated affinities of these groups are however difficult to explain. The Coelodendrida (Pl.121) are probably derived from the Concharida, and the Coelographida (Pl.126, 127, 128) from the Coelodendrida.
The geometrical fundamental form of the shell is in the majority of PHAEODARIA monaxonial, corresponding to the main axis of the enclosed central capsule; the astropyle of the latter, placed on the oral pole of the main axis, corresponds to the mouth of the shell in all Phaeogromia. In the Phaeosphaeria, where no pecuJiar shell-mouth is developed, the general fundamental form of the shell is usually homaxonial or spherical, often an endospherical polyhedron, rarely ellipsoidal or spindle-shaped (with prolonged main axis), or lenticular (with shortened main axis). The bivalved Phaeoconchia have usually either an amphithect shell (with the same fundamental form as the Ctenophora), or a dipleuric, bilaterally symmetrical shell (with a dorsal and a ventral valve, a right and a left parapyle). A small number of PHAEODARIA (mainly Circoporida) are remarkable on account of the regular polyhedral form of their shell, the geometrical axes of which resemble crystalline axes and are defined by regular radial tubes; as the octahedral Circoporus (Pl 117, fig. 6), the dodecahedral Circorrhegma (fig. 2), and the icosahedral Circogonia (fig. 1).
The siliceous or silicate bars, which compose the skeleton of the PHAEODARIA, are in the majority hollow tubes, filled up by jelly; in some other families, however, they are solid rods, as in the NASSELLARIA and SPUMELLARIA. Such usual lattice-work, composed of solid rods, occurs only in the families Sagosphaerida (Pl.108), Castanellida (Pl.113), and Concharida (Pls. 123, 124, 125). A quite peculiar structure, a diatomaceous tracery of extremely fine and regular hexagonal frames, distinguishes the Challengerida (Pl.99). The hollow cylindrical tubes, which are found in the other families, appear in three different forms, simple, articulate, and provided with an axial thread. Simple hollow tubes, which are neither articulate, nor provided with an axial thread, occur in the Cannorrhaphida (Pl.101) Aulacanthida (Pl.102-105), Coelodendrida (Pl.121, 122), and Coelographida (Pl.126, 127, 128). In all these families the hollow cylindrical tubes have a very thin wall and contain a wide cavity, filled only by jelly. The Orosphaerida (Pl.106, 107), differ in the reduction of the cavity, which becomes very narrow (often rudimentary or lost), whilst the walls of the tubes become extremely thickened and stratified, numerous concentric layers of silica being disposed one over the other. The hollow cylindrical tubes contain an axial filament, or a thin thread of silica, placed in its axis, in the families Aulosphaerida (Pl.109 - 111), Cannosphaerida (Pl.112), Circoporida (Pl.114-117), and Tuscarorida (Pl.100). Usually the axial filament is connected with the thin wall of the tube by numerous horizontal branches. A quite peculiar structure distinguishes the Medusettida (Pl.118 -120); their hollow tubes, extremely prolonged, are articulate owing to the presence of numerous, regular, equidistant transverse septa; these are pierced by a short tubule, similar to the siphon of the shells of Nautilus; this remarkable alveolar structure also occurs in the peripheral part of their shell-wall (and sometimes in the whole shell), numerous small polyhedral chambers or alveoles which communicate by small openings, being developed; they become easily filled with air in the dry shell (Pl.120, figs. 11-16).
The substance of the siliceous or siicated shell-wall is, in the majority of PHAEODARIA, homogeneous and structureless, as in the SPUMELLARIA and NASSELLARIA; but sometimes it acquires a peculiar structure. The thickened wall of the hollow tubes in the Orosphaerida and in several Aulacanthida (Pl.105, figs. 6-10) becomes distinctly stratified, concentric strata being disposed one over the other. A very remarkable structure, differing from that in all other Radiolaria, is found in the porcellanous shell of the Circoporida (Pl. 114 -117), and Tuscarorida (Pl.100). The thickened wall of the opaque shell is here composed of a peculiar silicated cement, which encloses numerous very thin and irregularly scattered needles (Pl. 115, figs. 6-9; Pl.116, fig. 3). Dry fragments of these shells, observed by a strong lens, appear finely punctulate, and probably air, entering into these fine porules of the cement, causes the white colour and the calcareous or porcellanous appearance of the opaque dry shell. Its surface is smooth in the Tuscarorida (Pl. 100), panelled in the Circoporida (Pl. 114 -117).
The hollow or solid spines, which arise from the shell of the PHAEODARIA,
exhibit an extraordinary variety and elegance in the production of different
branches, bristles, hairs, secondary spine, and thorns, hooks, anchor-threads,
pencils, spathillae, &c. These appendages are developed similarly to
those of many SPUMELLARIA, but exhibit a far greater variety and richness
in form. They are organs partly for protection, partly for retention of
food. They are much more interesting than in other Radiolaria.
|
|
 |
1 Ueber die Phaeodarien, eine ueue Gruppe
kieselschaliger mariner Rhizopoden,
Sitzungsb. med-nat. Gesellach. Jena,
December
12, 1579, pp. 3, 4.