{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

Nassellaria (inclusis Spyridinis), Ehrenberg, 1875.
Monopylea, Hertwig, 1879.
Monopylaria, Haeckel, 1881.
Cyrtida et Acanthodesmida, Haeckel, 1862.
Definition.Radiolaria with simple membrane of the central capsule, which is monaxon or bilateral, and bears on one pole of the main axis a porous area (porochora), forming the base of a peculiar intracapsular cone (podoconus). Extra-capsulum without phaeodium. Skeleton siliceous, very rarely wanting. Fundamental form originally monaxon, often dipleuric or bilateral.
The legion NASSELLARIA vel MONOPYLEA, in the extent here defined, was constituted in 1879 by Richard Hertwig in his work Der Organismus der Radiolarien (pp. 133-137). He gave to this large group the rank of an order, and united in it the two families Acanthodesmida and Cyrtida, which I had constituted first in 1862 in my Monograph (pp. 237, 265, 272); but he added, too, as a third family the Plagiacanthida, united by me with the former. In the first system of Ehrenberg (1847, loc. cit., pp. 53, 54), four families belonging to the MONOPYLEA were enumerated, the Halicalyptrina, Lithochytrina, Eucyrtidina, and Spyridina. He united the three former under the name "Polycystina solitaria," which he afterwards changed into NASSELLARIA (1875, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 157).
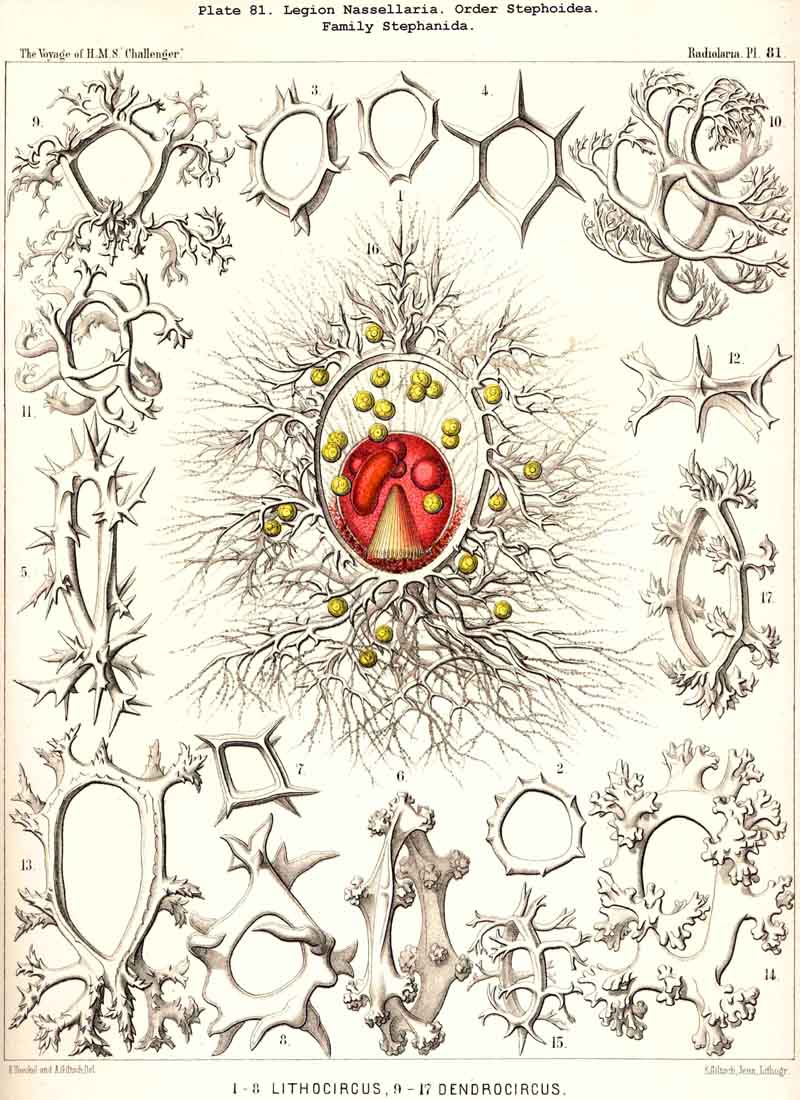
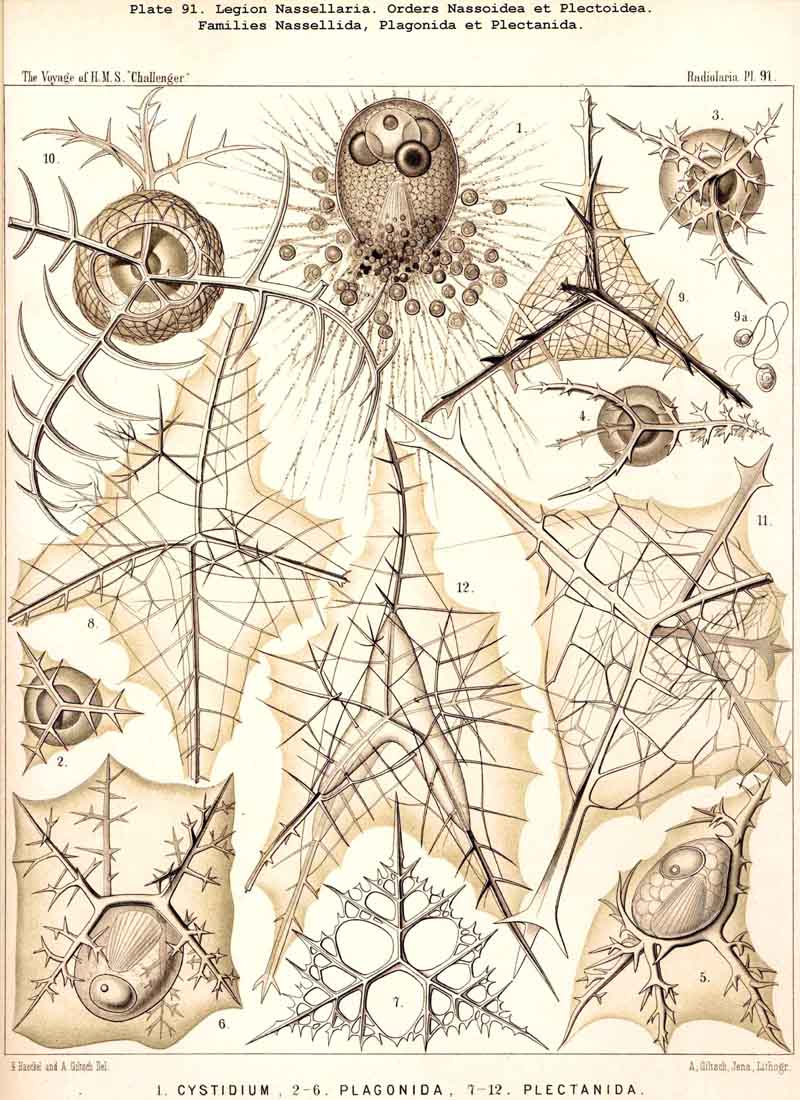
In my Monograph of the Radiolaria (1862, pp. 265-345) forty-four genera of NASSELLARIA were enumerated (six Acanthodesmida and thirty-eight Cyrtida), whilst the total number of genera in the whole class of Radiolaria at that time amounted to one hundred and thirteen. But owing to the astonishing number of new and interesting forms of this legion which I afterwards detected in the collection of the Challenger, in 1881 I distinguished in my Prodromus not less than three hundred and seventeen genera. These were disposed in five large main groups, retained in the present Report, with twenty-six families, viz., (1) Plectoidea (with three families), (2) Stephoidea (with four families), (3) Spyroidea (with four families), (4) Botryodea (with three families), and (5) Cyrtoidea (with twelve families). The first two groups have an incomplete or rudimentary skeleton, and may be united in the order Plectellaria, whilst the other three families possess a complete latticed shell, and may be united as Cyrtellaria. The former correspond to the Acanthodesmida, the latter to the Cyrtida in my Monograph.
The character common to all MONOPYLEA or NASSELLARIA, which separates them from all other Radiolaria, was first recognised by Richard Hertwig in 1879, and consists in the singular structure of the monaxonian central capsule, bearing on the basal pole a peculiar porous area or operculum, the "Porenfeld;" we call it shortly the "porochora." It represents a circular or elliptical porous plate on the basal pole of the vertical main axis of the central capsule, and bears a peculiar "podoconus" or "Pseudopodien-Kegel," a conical body of singular structure, protruding inside the membrane into the capsule. The pseudopodia arising from this peculiar "podoconus" proceed from the capsule piercing the "porochora," whilst the other parts of the capsule are not perforated.
The MONOPYLEA are therefore "Merotrypasta," like the following fourth legion, the PHAEODARIA or CANNOPYLEA. But in these latter we find on the basal pole of the monaxonian capsule only one single large main opening, prolonged into a peculiar tube, and there is no trace of the typical "podoconus," characteristic of all NASSELLARIA. The latter agree, however, with the former in the possession of a basal opening, serving for the emission of the pseudopodia, and in the monaxonian fundamental form, arising from this structure. Therefore these two legions of "Merotrypasta" exhibit a wider divergence from the ACANTHARIA and SPUMELLARIA, the two legions of "Holotrypasta," in which the central capsule is everywhere perforated by innumerable small pores (compare above, pp. 5, 6, 716).
The Skeleton in all NASSELLARIA consists either of pure silica, or of a peculiar silicate; never of acanthin (as in all ACANTHARIA). The siliceous bars and beams constituting it are invariably solid (as also in the SPUMELLARIA); never hollow (as in the PHAEODARIA). In the small family of Nassellida alone (with the two genera Cystidium and Nassella) the skeleton is entirely absent. In all other genera of NASSELLARIA the siliceous skeleton is more or less developed ; imperfect, or quite rudimentary in the Plectellaria (Plectoidea and Stephoidea), but perfect and forming a regular lattice-shell in the Cyrtellaria (Spyroidea, Botryodea, and Cyrtoidea). The different forms of this skeleton exhibit an extraordinary variety, but may be reduced to a few very simple fundamental forms, or even to a single, most simple original form. The comparative morphology is more interesting, but also more difficult to understand than in any other Radiolaria.
The geometrical fundamental form of the skeleton is in all NASSELLARIA monaxonial, the vertical main axis of the body, which is already indicated by the axis of the central capsule with two different poles, being also expressed constantly in the form of the skeleton. The lower or basal pole of the latter always exhibits, a different shape from that of the upper or apical pole. This difference is so striking in nearly all MONOPYLEA, that the two poles may be determined on the first view.
In the great majority of NASSELLARIA not only is the monaxonial fundamental form expressed, but also the dipleuric or bilateral, so that the symmetrical halves of the body may be easily determined; the right and left halves exhibit the same symmetry as in the vertebrates, so that we may distinguish an anterior ventral and a posterior dorsal face of the body. The whole form is in this case determined by three dimensive axes, perpendicular to one another, two of which are heteropolar, the third is homoeopolar. The apical pole of the vertical main axis (principal or longitudinal axis) is different from the basal pole. The ventral pole of the horizontal sagittal axis (or dorso-ventral axis) is different from the dorsal pole. The right pole of the horizontal transverse axis (lateral or frontal axis) is equal to the left pole. Therefore the sagittal or median plane of the body (in which the principal and the sagittal axis are crossed) divides it into symmetrical equal halves and is perpendicular to the frontal axis.
Three different original elements of structure are recognisable in the majority of NASSELLARIA, viz., (1) a vertical simple ring, the primary or sagittal ring, placed vertically in the sagittal plane and enclosing the median plane of the central capsule; (2) a basal tripod, composed of three diverging radial rods, which are united on the basal pole of the central capsule and are either expanded horizontally or descend; (3) an ovate or subspherical, simple lattice-shell, the cephalis or capitulum, which surrounds the central capsule and exhihits a peculiar structure on its basal pole.
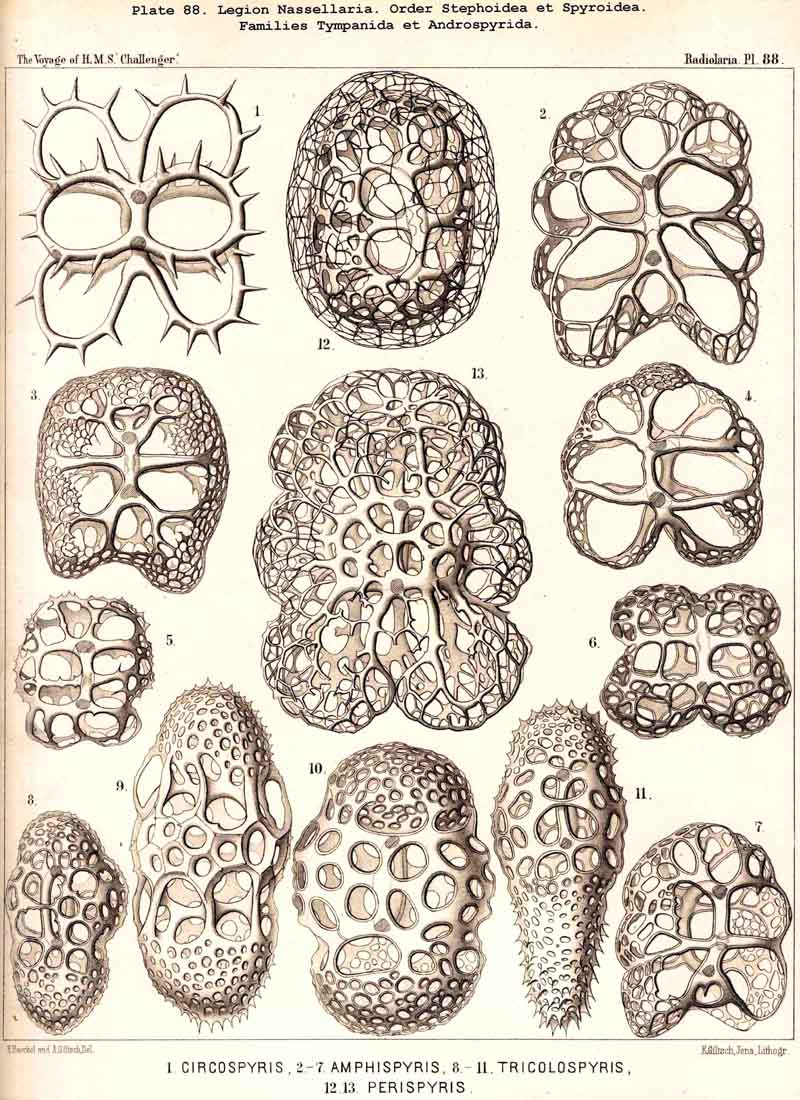
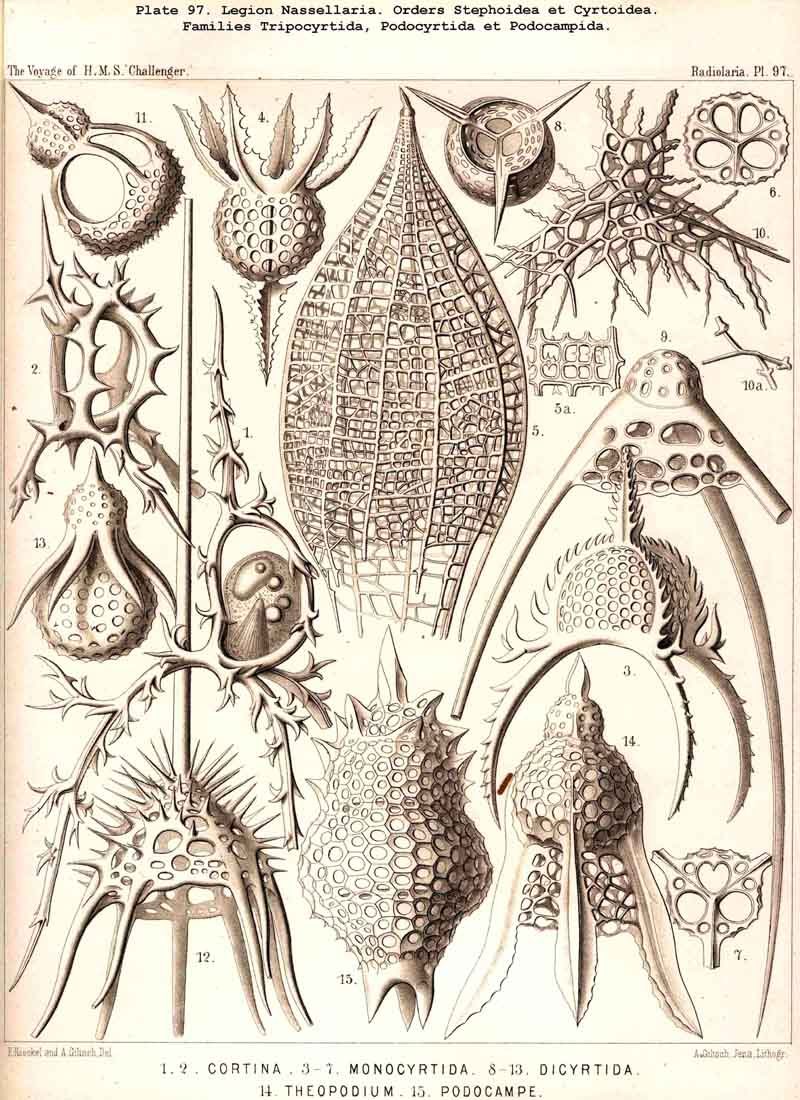
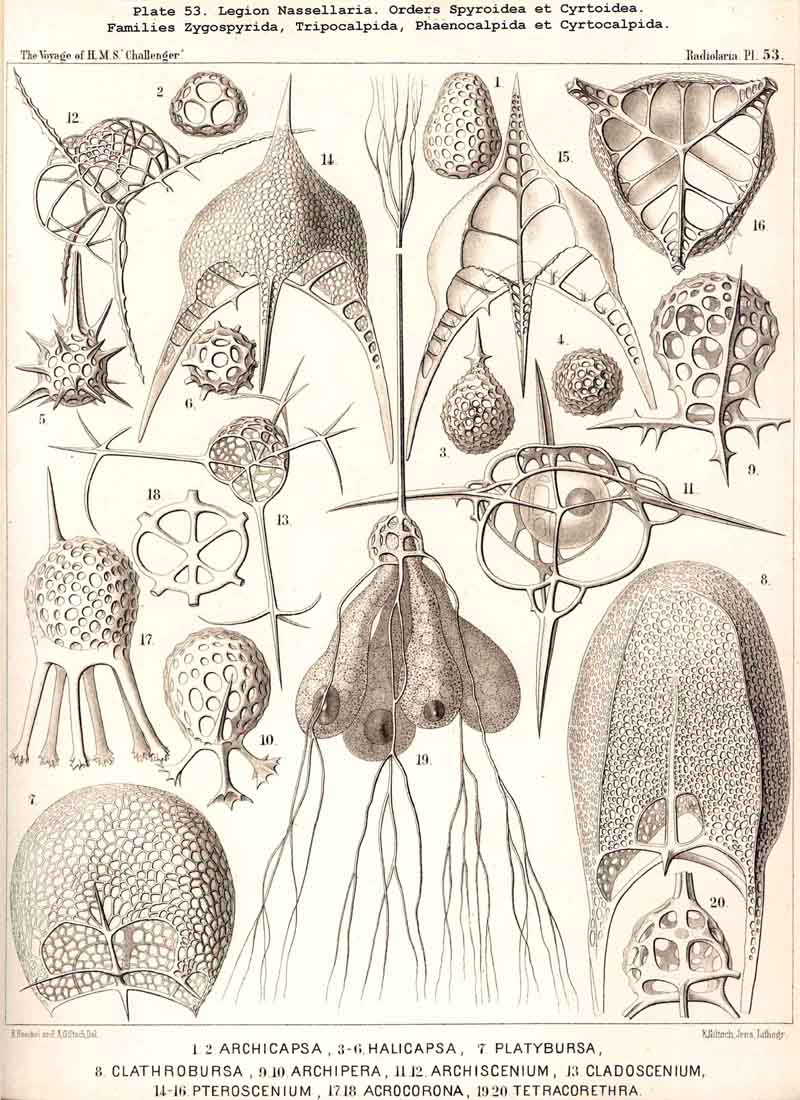
These three important original elements of structurethe sagittal ring, the basal tripod, and the latticed cephalisare so united in the majority of NASSELLARIA that the cephalis rests upon the tripod and includes the sagittal ring wholly or partially. The simplest realisation of this typical union is afforded by the Archiperida and Tripospyrida, and these may be derived from the simpler important Stephanid Cortina (Pl. 88, fig. 9; Pl. 92, fig. 21; Pl. 97, fig. 1). In this and in all other tripodal NASSELLARIA, the three basal rods or the "cortinar feet" are constantly so arranged that an odd or posterior rod, the "caudal foot" (c) is opposed to the two anterior paired rods, the "pectoral feet" (one right, p", and one left, p'). The caudal foot lies in the sagittal plane, and is prolonged upwards into the dorsal rod of the sagittal ring (b), and over this in a free ascending spine, the "apical horn" (a). The curved ventral rod of the ring (r) is united above with the base of the apical horn, below with the common centre of the tripod or the "cortinar centrum." The characteristic position of the central capsule in this skeleton of Cortina is such that its basal pole (with the porochora) rests upon the centre of the tripod, whilst its sagittal perimeter is separated from the surrounding ring by the calymma; the numerous pseudopodia arising from its base diverge downwards and are supported and protected by the three basal feet of the tripod (Pl. 97, fig. 1). Compare also Pls. 51, 53, 84, 95, 98.
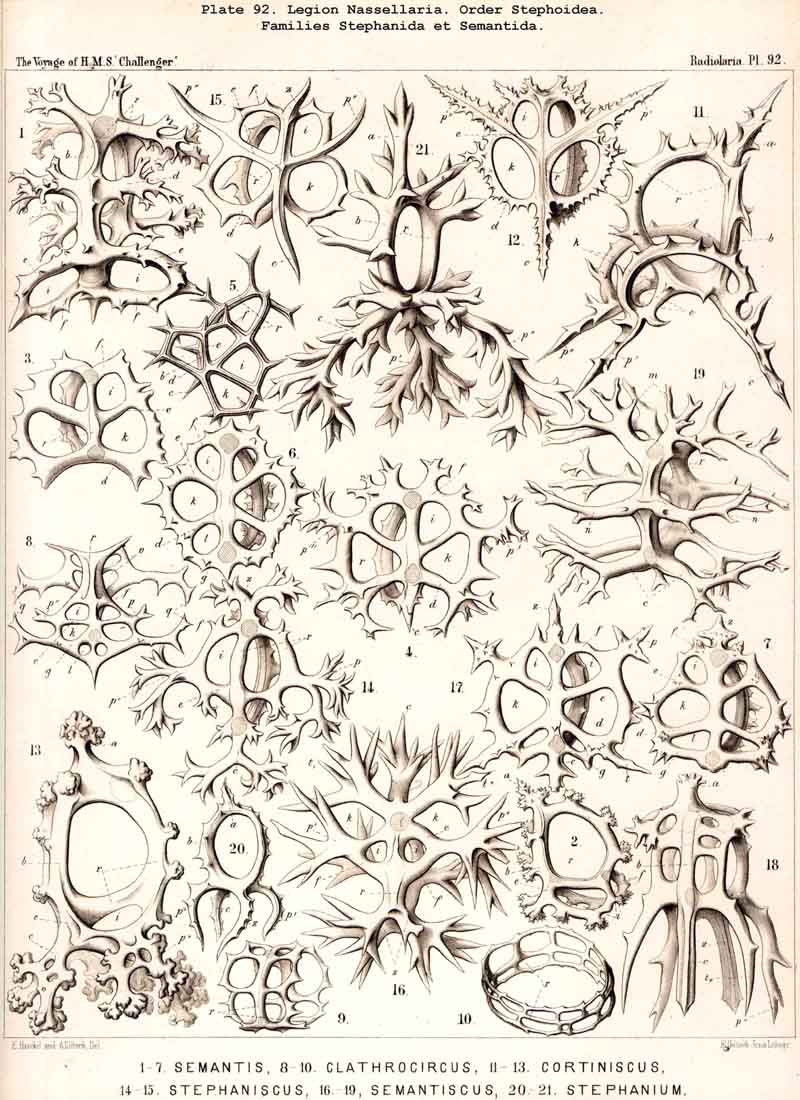
The typical skeleton of Cortina, a tripodal ring, becomes more developed in the Semantid Cortiniscus, in which the basal parts of the three diverging feet are united by a second horizontal ring, the cortinar or basal ring (Pl. 92, figs. 11-13). The pores between the former and the latter, or the "cortinar pores," may be regarded as the first beginning of the lattice-plate, composing the "cephalis" or the simple primordial shell in the Archiperida and Tripospyrida, and transmitted from these by heredity to the great majority of NASSELLARIA.
The "cephalis or capitulum" (the "Köpfchen" of the German authors) is therefore the most important part of the skeleton in all Cyrtellaria, or in all NASSELLARIA possessing a complete lattice-shell. In the Plectelaria, however, or in those NASSELLARIA which do not possess a complete fenestrated shell, the "cephalis" is either imperfect or totally wanting. The cephalis surrounds the enclosed central capsule on all sides in the form of an ovate or subspherical lattice-shell, and is separated from it only by the jelly-like calymma. The sagittal ring is either enclosed in the wall of the cephalis (wholly or partially), separating its two lateral halves, or it is enclosed in the cavity of the cephalis and connected with its sagittal perimeter by short beams. The base of the cephalis (with the cortinar plate) often rests immediately upon the centre of the tripod; in the majority of NASSELLARIA, however, this near relation is altered by reason of later changes and secondary modifications.
The number of various forms, developed from these simple original structural elements of the skeleton, is astonishing, and there are described more than three-hundred genera and nearly two thousand species of this legion in the following pages. This large number may be easily increased by subsequent observers. Since in all these MONOPYLEA the characteristic structure of the central capsule is identical, and, also the structural elements of the siliceous skeleton are similar, it is very probable that they may have arisen from a single common stock. But it is very difficult (and at present impossible) to explain a natural monophyletic system of this large legion. The greatest difficulty is presented by the fact that the three structural elements mentioned above, viz., the sagittal ring, the basal tripod, and the latticed cephalis, are not constantly united, but each alone may also constitute the skeleton by itself. In this respect the following seven cases are possible, and are actually realised.
A. The skeleton is composed of the sagittal ring only and of its spiny appendages, without basal tripod and without latticed cephalis. This is the case in the majority of Stephoidea (Stephanida, Semantida, Coronida, and Tympanida).
B. The skeleton is composed of a basal tripod only (Plagiacantha), or of a tripod in the centre of which arises a vertical apical horn (Plagoniscus), and often of an irregular framework, arising from the rods of the tripod; but there is neither a trace of a sagittal ring nor a complete latticed cephalis. This is the case in the remarkable suborder Plectoidea (Plagonida and Plectanida).
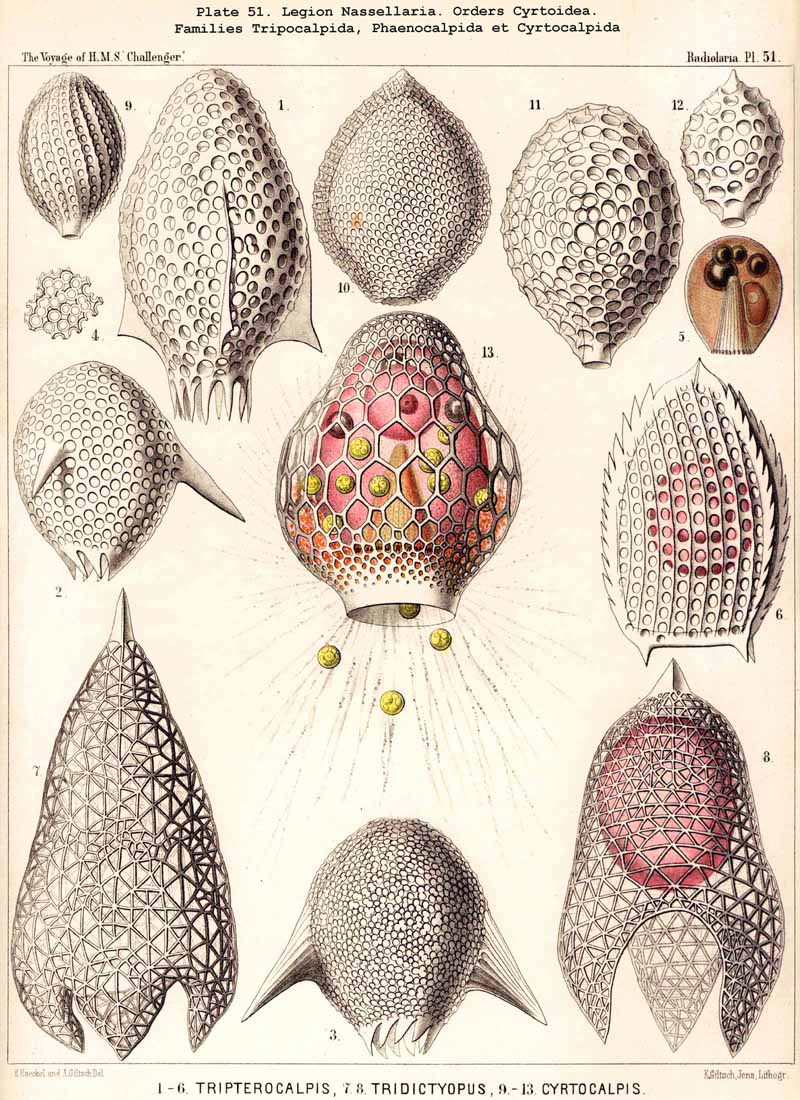
C. The skeleton is composed of a latticed cephalis only, a simple ovate or sub- spherical fenestrated shell, which encloses the monopylean central capsule; there is no trace of a sagittal ring nor of a basal tripod. This is the case in the remarkable family of Cyrtocalpida (Archicorida and Archicapsida), in numerous Botryodea and in other Cyrtellaria.
D. The skeleton is composed of a sagittal ring and a basal tripod, without latticed cephalis. This is the case in a few, but very important forms of Stephoidea: Cortina, Stephanium, Cortiniscus, Stephaniscus, Podocoronis, and some allied genera.
E. The skeleton is composed of a sagittal ring and a latticed cephalis, but without basal tripod. This is the case in numerous Cyrtellaria, in the Circospyrida (or Zygospyrida apoda: Dictyospyris, Circospyris) and some other Spyroidea; and in a large number of Botryodea and Cyrtoidea eradiata (a part of the Sethocyrtida, Theocyrtida, Lithocampida, and others).
F. The shell is composed of a basal tripod and a latticed cephalis, but without any trace of the sagittal ring. This is the case in numerous Cyrtoidea triradiata and multiradiata, and perhaps in the majority of the following families Tripocalpida, Tripocyrtida, Podocyrtida, and Podocampida.
G. The shell is composed of all three above-mentioned elements, of a sagittal ring, a basal tripod, and a latticed cephalis. This is the case in the great majority of Spyroidea (with a few exceptions only), and perhaps also in the majority of Cyrtoidea.
The survey of these seven groups, A to G, each of which is represented by numerous living forms, shows clearly how difficult and complicated the morphology and phylogeny of the numerous NASSELLARIA must be. For all possible combinations of the three original structural elements are realised abundantly, and in such complicated relations, and so intermingled in the different orders and families, that it seems nearly hopeless to answer the question of their true origin. The identity in the structure of the central capsule, however, in all these MONOPYLEA, makes it probable that they have all arisen originally from the skeletonless Nassellida (Cystidium, Nassella), either in a monophyletic or in a polyphyletic way. In this respect the following phylogenetical hypotheses are possible.
1. Monophyletic hypothesis, deriving all NASSELLARIA from a simple sagittal ring (Archicircus, Lithocircus, &c., Pl. 81). The groups A, D, E, and G may be derived easily from such a ring, but the groups B, C, and F only by means of the hypothesis that the original ring may be completely reduced and finally lost. This hypothesis was stated by me in the years 1877 to 1879, when I had got the first general survey of the astonishing number of new NASSELLARIA in the Challenger collection, and as I had found the sagittal ring in the majority of them. This, my former hypothesis, is mentioned by Richard Hertwig (1879, loc. cit., pp. 68, 126). It was afterwards supported with particular energy by O. Bütschli (1882, Zeitschr. für wiss. Zool., Bd. XXXVI.).
2. Monophyletic hypothesis, deriving all NASSELLARIA from a basal tripod (Triplagia, Plagoniscus, &c., Pl. 91). The groups B, D, F, and G, all triradiate, may be derived easily from such a tripod; but the groups A, C, and E only by means of the hypothesis that the original tripod may be completely reduced and finally lost. This hypothesis was employed in 1881 in my Prodromus, since I had convinced myself that the "triradial structure" is prevalent in the great majority of NASSELLARIA, and is perhaps more important than the sagittal ring.
3. Monophyletic hypothesis, deriving all NASSELLARIA from a latticed cephalis, a simple ovate or subspherical fenestrated shell without ring and tripod (Cyrtocalpis, Archicapsa, &c.). The groups C, E, F, and G may be derived easily from such a cephalis, but the groups A, B, and D only by means of the hypothesis that the sagittal ring as well as the basal tripod may remain as the last remnants of a reduced cephalis. This hypothesis was given in 1862 in my Monograph, where I constructed the first pedigree of Radiolaria (p. 234). I there derived all the Cyrtida from the Sphaeroidea (Cyrtidosphaera), supposing that Cyrtocalpis and some other Monocyrtida may form a direct phylogenetical passage from the Sphaeroidea to the Cyrtoidea.
4. Polyphyletic hypotheses, deriving the different groups of NASSELLARIA from different skeletonless Nassellida, by development of simple siliceous skeletons in different ways. Among the numerous polyphyletic hypotheses which are possible, one of the simplest would be the supposition that three different fundamental forms of skeleton may have arisen independently one from another: (1) a simple sagittal ring as original form of the Stephoidea and Spyroidea (A); (2) a simple basal tripod as original form of the Plectoidea (B); (3) a simple latticed cephalis as original form of the Botryodea and Cyrtoidea (C). This triphyletic hypothesis is supported by R. Hertwig (1879, loc. cit., p. 136); he assumes that the original skeletonless Nassellida (Cystidium) have produced three different branches, his "Acanthodesmida" (=Stephoidea and Spyroidea) with a primary ring, his "Plagiacanthida" (=Plectoidea) with a primary tripod, and his Cyrtida (Botryodea and Cyrtoidea) with a primary cephalis. This hypothesis seems rather probable on the first view; but it meets with the greatest difficulties in view of the fact that these three original elements of the skeleton are more or less evidently combined in the great majority of NASSELLARIA. The greatest difficulty arises from the fact that often among very similar and closely allied forms the first exhibits all three elements (A, B, C) combined, whilst the second has a combination of A and B, the third of B aud C, the fourth of A and C; and there are other forms, very similar to the former, in which one element only is recognisable. Another difficulty arises from the fact that the intimate structure of the cephalis in the majority of Cyrtellaria is not perfectly known, and often exhibits structures which are difficult to explain with regard to the three elements A, B, C. Under these circumstances further researches on the numerous imperfectly known NASSELLARIA are required, and chiefly accurate observations on their more minute structure and on their important ontogeny.
We divide the immense legion of NASSELLARIA into two large orders, the
Plectellaria without complete lattice-shell, and the Cyrtellaria with a
complete lattice-shell or a "cephalis," including the central capsule;
the latter, of course, have arisen from the former. The Plectellaria comprise
three different suborders, the Nassoidea (without skeleton), the Plectoidea
(with a tripodal skeleton, without ring), and the Stephoidea (with a primary
sagittal ring, with or without tripod). The Cyrtellaria again also comprise
three different suborders, the Spyroidea (with biocular cephalis and a
sagittal constriction), the Botryodea (with multilocular and lobate cephalis,
exhibiting two or more constrictions and three or more lobes), and the
Cyrtoidea (with simple, unilocular cephalis, without constriction).
|
|