(§§ 218-225.)
218. Motion.-In addition to the internal movements which appear generally in the unicellular Radiolaria and have already been mentioned as plasmatic currents m treating of the circulation (§§ 207-209), two different groups of external motor phenomena are to be observed in this class: first, the contraction of individual parts, which brings about modifications of form (§ 220), and secondly, voluntary or reflex locomotion of the whole body (§ 220). These movements are partly due to changes in form of undifferentiated plasmatic threads or sarcode filaments, partly to the actual contraction of differentiated filaments which are comparable to muscle fibrillae, and must therefore be distinguished as myophanes. In addition to this, endosmose and exosmose probably play an important part in some of the locomotive phenomena, but nothing is yet certainly known regarding these osmotic processes. We are at present equally ignorant whether all the movements of the Radiolaria are simply reflex (direct consequences of irritation) or whether they are in part truly spontaneous.
219. Suspension. -From direct observation of living Radiolaria, as well as from deductive reasoning, based upon their morphology (and especially their promorphobogy, §§ 17-50), the conclusion appears justified that all Protista of this class in their normal condition float suspended in the sea-water, either at the surface or at a definite depth. A necessary condition of this hydrostatic suspension is that the specific gravity of the Radiolarian organism must be equal to, or but slightly greater than that of sea-water. The increase in specific gravity brought about by the production of the siliceous skeleton, is compensated by the lighter fatty gbobules, and partly perhaps by the calymma, especially when the latter contains vacuoles or alveoles. The fluid or jelly contained in the latter appears to be for the most part lighter than sea-water (containing no salt, or only a very small quantity ?). But if the specific gravity of the whole body should be generally (or perhaps always) slightly greater than that of sea-water, then the organism would be prevented from sinking, partly by the increased friction, due to the radiating pseudopodia and the radial spines usually present, and partly perhaps by active (if only feeble) movements of the pseudopodia.
220. Locomotion.-Active locomotion of the whole body, which is very probably to be regarded as voluntary, occurs in the Radiolaria in three different modes ; (1) the vibratile movement of the flagellate swarm-spores ; (2) the swimming of the floating organisms ; (3) the slow creeping of those which rest accidentally upon the bottom. The vibratile movement of the swarm-spores is the result of active sinuous oscillation of the single or multiple flagellum, and is not essentially different from that of ordinary flagellate Infusoria (see note A). Of the active swimming of mature Radiolaria, only that form is known which is vertical in direction and causes the sinking and rising in the sea-water. This is probably, for the most part (perhaps exclusively), due to increase or diminution in the specific gravity, which is perhaps brought about by the retraction or protrusion of the pseudopodia ; slow, oscillating, sinuous motions of these organs have been directly observed to take place (though very slowly) in suspended living Radiolaria. The most important hydrostatic organ is probably the calymma, by the contraction of which the specific gravity is increased, while it is diminished by its expansion ; the contraction is probably brought about by active contraction of the sarcodictyum, and is connected with exosmosis, while the expansion is probably due to the elasticity of the calymma and the inception of water by endosmosis. In the Acanthometra (§ 96) the peculiar myophriscs appear to be charged with the duty of distending the gelatinous envelope, and thus diminishing the specific gravity ; the latter increases again when the myophriscs are relaxed, and the calymma contracts by virtue of its own elasticity (see note B). The slow creeping locomotion exhibited by Radiolaria on a glass slide under the microscope, does not differ from that of the Thalamophora (Monothalamia and Polythalamia), but can only occur normally when the animal accidentally comes into contact with a solid surface or sinks to the bottom of the sea. Whether this actually occurs periodically is not known (see note C). The slow or gliding locomotion exhibited by creeping Monozoa on a glass slide is due to muscle-like contractions of bundles of pseudopodia, just as in the case of the social central capsules of Polyzoa, which live together in the same coenobium and are able to move within their common calymma sometimes centrifugally to its surface, sometimes towards the centre where they aggregate into a roundish mass (see note ID).
A. Regarding the movement of the flagella in mature swarm-spores compare L. N. 22, p. 375 ; L. N. 26, pp. 31, 35; L. N. 41, p. 452, and L. N. 52, p. 170.
B. On the active vertical swimming movements of mature Radiolaria, especially the cause of sinking and rising, see 1. N. 16, p. 134 ; 1. N. 41, p. 443, and L. N. 52, pp. 97-102.
C. On the active horizontal creeping movements of mature Radiolaria on a firm ground, compare L N. 12, p. 10, and L. N. 16, pp. 132-134.
D. Regarding the motion of social central capsules within the same ccenobium and the changes thus brought about in the structure of the calymma, see 1. N. 16, pp. 119-127, and L. N. 52, pp. 75-82.
221. Contraction.-Motions, which are due to the contraction of individual portions and cause changes in volume or form, have been partly already spoken of under the head of locomotion (§ 220) and are partly connected with other functions. Examples may be seen in the contraction of the central capsule and of the calymma. A certain contraction of the central capsule is probably brought about by the myophanes, which arise by differentiation of the endoplasm and hence may assume different forms in the four legions. In the SPUMELLARIA, where numerous radial fibrillae run from the central nucleus to the capsule membrane (§ 77), the endoplasm is probably driven out evenly through all the pores of the capsule membrane by their simultaneous contraction, and hence the volume of the capsule is diminished in all directions. The ACANTHARIA probably behave similarly, but are different, inasmuch as the number of their contractile radial fibrillae is less, and special axial threads (§ 78) are already differentiated. In the NASSELLARIA it is probable that owing to the contraction of the divergent myophane fibrillae in the podoconus the vertical axis of the latter is shortened, the opercular rods of the porochora are lifted, and the endoplasm driven out of its pores, so that the volume of the monaxon central capsule is diminished (§ 79). In the PHAEODARIA the same result is probably brought about by the contraction of the cortical myophane fibrillae, which run meridionally along the inside of the capsule membrane from the apical to the basal pole of the vertical main axis, where they are inserted into the periphery of the astropyle ; since the volume of the capsule is diminished by their contraction (their spheroidal figure becoming more nearly spherical) the endoplasm will be driven out through the proboscis of the astropyle. Whilst these contractions of the central capsule are largely due to differentiated muscle-like threads of endoplasm (myophanes), this appears to be but rarely the case with the contractions of the extracapsulum (e.g., the myophriscs of the Acanthometra, § 96). Most of the phenomena of contraction which can be observed in the calymma and pseudopodia depend upon exoplasmatic currents (§ 209).
222. Protection.-Of the utmost importance, both for the physiology and for the morphology of the Radiolaria are their manifold protective functions, which we now consider under the heading "protection." From the physiological point of view the consideration of the exposed situation in which the delicate, free-swimming Radiolarian organism lives, and the numerous dangers which beset it in the struggle for existence, would lead a priori to the expectation, that many special protective adaptations would be developed by natural selection. On the other hand, morphological experience shows us that this latter has been in action for immeasurable periods, and has gradually produced an abundance of the most remarkable protective modifications. Examples of these may be found in the formation of the voluminous calymma, as a gelatinous protective covering for the central capsule, and further, the formation of the capsule membrane itself, which separates the generative contents of the central capsule from the nutritive exoplasm. The phosphorescence of the central capsule, too (§ 223), may be regarded as a useful protective arrangement ; as also the radiating of the numerous pseudopodia in all directions from the surface of the calymma ; for they are of great significance to the well-being of the organism, both as sensory organs and as prehensile organs. By far the most important and most varied means for the actual defence of the soft body is to be seen in the endless modifications of the skeleton ; first, in the production of the enclosing lattice-shells and projecting radial spines, but especially also in the very varied structure of the individual parts of the skeleton, and in the special differentiation of the small appendicular organs which grow out from it (hairs, thorns, spines, scales, spathillae, anchors, &c.). Finally "mimicry" possesses a considerable significance among the different forms of adaptation which are to be observed in this class.
223. Phosphorescence.-Many Radiolarians shine in the dark, and their phosphorescence presents the same phenomena as that of other luminous marine organisms ; it is increased by mechanical and chemical irritation, or renewed if already extinguished. The light is sometimes greenish, sometimes yellowish, and appears generally (if not always) to radiate from the intracapsular fatty spheres (§ 73). Thus these latter unite several functions, inasmuch as they serve, firstly, as reserve stores of nutriment, secondly, as hydrostatic apparatus, and thirdly, as luminous organs for the protection of the Radiolaria ; probably the light acts by frightening other animals, for the phosphorescent animals are provided with spines, nettle-cells, poison glands or other defensive weapons. The production of the light depends probably, as in other phosphorescent organisms, upon the slow oxidation of the fat-globules, which combine with active oxygen in the presence of alkalis. Phosphorescence is very likely widely distributed among the Radiolaria.
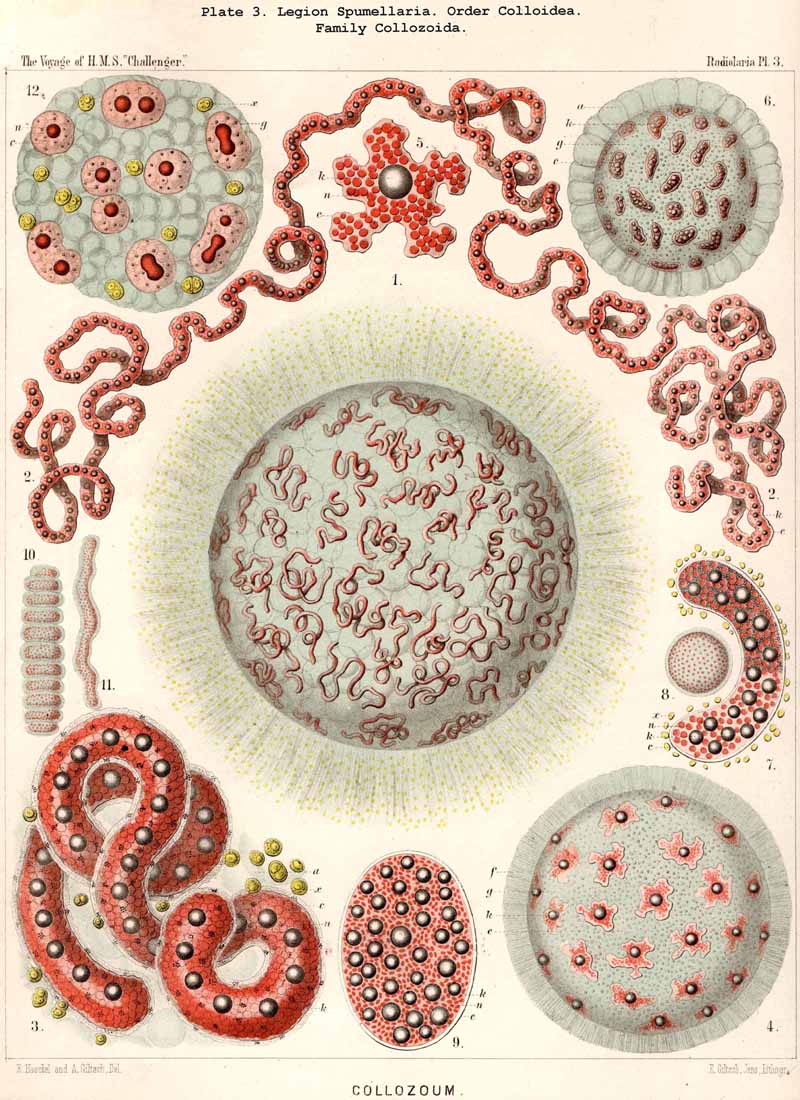
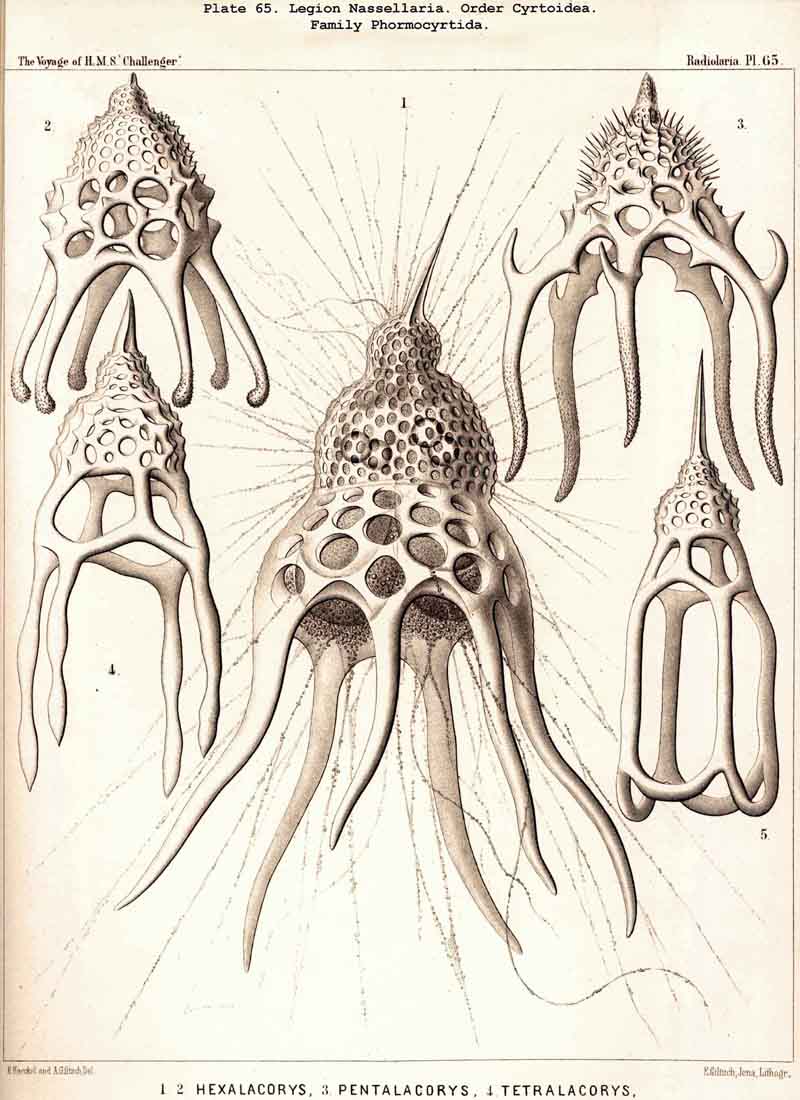
The shining of the Radiolaria in the dark has been noticed by the earliest observers of the class (see L. N. 1, p. 163, L. N. 16, p. 2, and L. N. 52, pp. 136-139). In the winter of 1859 I observed the production of light in the case of many monozootic and polyzootic Radiolaria, but inadvertently omitted to record the fact in my Monograph. I made more accurate observations in the winter of 1866 at Lanzerote in the Canary Islands, and convinced myself that the light emanates from the central capsule, and in particular from the fat-globules contained in it. In most Polycyttaria (both Collosphaerida and Sphaerozoida), when each central capsule contains a large central oil-globule the light radiates from it. In Collozoumm serpentinum(Pl. 3, figs. 2, 3) each cylindrical central capsule contains a row of luminous spherules like a string of beads. In Alacorys friderici (Pl. 65, fig. 1) the four-lobed central capsule contains four shining points. Karl Brandt has recently made more detailed communication on this point (L N. 52, p. 137).
224. Sensation.-The general irritability which we ascribe to all organisms, and as the basis of which we regard the protoplasm, remains at an inferior stage of development in the Radiolaria. For although they are subject to various stimuli, and certainly possess a power of discrimination, special sensory organs are not differentiated ; the peripheral portions of the protoplasm, and especially the pseudopodia, rather act both as organs of the different kinds of sensation and various modes of motion. That different Radiolaria have attained different degrees of development in this respect may be seen partly by direct observation of the reaction of the living organism towards various stimuli, and partly by the comparison of the different conditions of existence under which Radiolarians exist, both in the most various depths of the ocean and in all climatic zones (see note A). In general the Radiolaria seem to be sensitive to the following stimuli ; (1) pressure (see note B) ; (2) temperature (see note C) ; (3) light (see note D) ; (4) chemical composition of the sea-water (see note E). The reaction towards these stimuli, corresponding to the sensation of pleasure or dislike which they call forth, is shown in various forms of motion of the protoplasm, changes in the currents in it, contraction of the central capsule, changes in the size, position, and form of the pseudopodia, changes in the volume of the calymma (by the evacuation of water), &c. Among the sensory functions of the Radiolaria must be especially mentioned their remarkably developed perception of hydrostatic equilibrium (see note F), as well as their perception of distances, so clearly shown in the production of equal lattice-meshes and other regularly formed skeletal structures (see note G).
A. I can add but little to the communication which I made twenty-four years ago regarding sensation in the Radiolaria (L. N. 16, pp. 128-131). The most important point would be the great difference in irritability which must obtain between the pelagic, zonarial and abyssal Radiolaria, which may be assumed from a consideration of their very different conditions of existence as re gards pressure, light, warmth, nutrition, &c. It is natural to suppose that the numerous abyssal Radiolaria, discovered by the Challenger, which live at great depths (2000 to 4500 fathoms) in complete darkness, in icy cold and under an enormous pressure, must have quite different sensations of pleasure from their pelagic relatives which live at the surface of the sea under an equatorial sun. Karl Brandt has recently added much to our knowledge regarding the special action of different vital conditions upon the various Polycyttaria and the degrees of their irritability (L. N. 52, pp. 113-132).
B. Regarding the sensation of pressure or sensation of touch of the Radiolaria and the various degrees of their mechanical irritability, see L. N. 16, p. 129 ; L. N. 41, p. 464.
C. Regarding the sensation of warmth or temperature-sense and its dependence upon different climatic relations, see L. N. 16, p. 129; L. N. 52, pp. 114-129.
D. Regarding the sensation of light, compare L. N. 16, p. 128 ; L. N. 42, p. 304; L. N. 52, pp. 102-104, 114.
E. Regarding the sense of taste of the Radiolaria or their peculiar sensitiveness towards the different chemical composition of the water, change in its salinity, presence of organic impurities, see L. N. 16, p. 130; L. N. 52, pp. 103, 113. This chemical irritability seems to be the most highly developed sense in the Radiolaria, even more so than their mechanical irritability.
F. The perception of hydrostatic equilibrium among the Radiolaria is immediately visible from the position which their bodies, floating freely in the water, assume spontaneously, and from the symmetrical development of the skeleton, which by its gravitation necessitates a definite position. It may be assumed that the development of the various geometrical ground forms which correspond to a definite position of equilibrium, is the result of this particular kind of perception (compare §§ 40-45).
G. The plastic perception of distance of the pseudopodia is shown by the symmetry with which the forms composing the regular skeletal structures (e.g., the ordinary lattice-spheres with regular hexagonal meshes, the radial spines with equidistant branches) are excreted from the exoplasm. Both this form of sensation and the one first mentioned (note F) have hitherto received scarcely any attention, but are deserving of a thorough physiological investigation.
225. The Cell-Soul (Zellseele).-The common central vital principle, commonly called the "soul," which is considered to be the regulator of all vital functions, appears in the Radiolaria as in other Protista in its simplest form, as the cell-soul. By the continual activity of this central "psyche" all vital functions are maintained in unbroken action, and in uniform correlation. It is also probable that by it the stimulations which the peripheral portions of the cell receive from the outer world are first transmitted into true sensation, and that, on the other hand, the volition, which alone calls forth spontaneous movements, proceeds from it. The central capsule is most likely the sole organ of this cell-soul or central psychic organ, and the active portion may be either the endoplasm or the nucleus, or both. The central capsule may thus (apart from its function as a sporangium, § 215) be regarded as a simple ganglion cell, physiologically comparable to the nervous centre of the higher animals, whilst the exoplasm (sarcomatrix and pseudopodia) are to be compared to the peripheral nervous system and sense organs of the latter. The great simplicity of the functions of the cell-soul which appear in the Radiolaria, and the intimate connection of their different psychic activities, give to these unicellular Protista a special significance for the comprehension of the monistic elements of a natural psychology.
Regarding the theory of the cell-soul as the only psychological theory which is able to explain naturally the true nature of the life of the soul in all organisms as well as in man, see my address on cell-souls and soul-cells ("Zellseelen und Seelenzellen") in Gesammelte populare Vortrage aus dem Gebiete der Entwickelungslehre, Heft 1, p. 143 ; Bonn, 1878.
{kind=link}
{kind=link}