CHAPTER VII.VEGETATIVE FUNCTIONS.
(§§ 201-217.)
201. Mechanism of the Functions.-The vital phenomena of the Radiolaria are dependent upon the mechanical functions of their unicellular body, and like those of all other organisms, are to be referred to physical and chemical natural laws. All processes which appear in the life of the Radiolaria are, therefore, ultimately to be explained by the attraction and repulsion of the smallest particles, which compose the different portions of their unicellular body ; and the sensation of pleasure or the opposite is in its turn the exciting cause of these elementary movements. Many adaptive arrangements in the Radiolarian organism may produce the appearance of being the premeditated result of causes working towards an end (" zweckthältig," causae finales), but as opposed to this deceptive appearance it must here be expressly stated that these may be recognised in accordance with the developmental theory as the necessary consequence of mechanical causes (causae efficientes).
Our physiological acquaintance with the Radiolaria has by no
means progressed so far as our morphological, so that the incomplete
communications which are placed here for the sake of complete ness must
be regarded merely as preliminary fragments, not as fully elaborated results.
Since my recent investigations have been mainly in the direction of morphology,
I can add but little to the physiological conclusions, which I stated at
length in my monograph twenty-four years ago (L. N. 16, pp. 127-165). Recently
the vegetative physiology of the Radiolaria has been much advanced by the
recognition of the symbiosis with the Xanthellae (§ 205, L. N. 22,
39, 42). In addition Karl Brandt has recently (1885) published several
important contributions to the physiology of the Polycyttaria or Sphaerozoea
(L. N. 52).
202. Distribution of Functions.-The distribution of the functions among the various parts of the unicellular organism of the Radiolaria corresponds directly to their anatomical composition, so that physiologically as well as morphobogically the central capsule and the extracapsulum appear as the two coordinated main components. On the one hand the central capsule with its endoplasm and enclosed nucleus is the central organ of the "cell-soul" (Zellseele), the unit regulating its animal and vegetative functions, and the special organ of reproduction and inheritance. The extracapsulum forms, on the other hand, by its calymma the protective envelope of the central capsule, the support of the soft pseudopodia and the substratum of the skeleton ; the calymma acts also as a hydrostatic apparatus, whilst the radiating pseudopodia are of the greatest importance both as organs of nutrition and adaptation, as well as of motion and sensation (§ 15). If, however, the vital functions as a whole be divided in accordance with the usual convention into the two great groups of vegetative (nutrition and reproduction) and animal (motion and sensation), then the central capsule would be mainly the organ of reproduction and sensation, and the extracapsulum the organ of nutrition and motion.
The numerous separate vital phenomena, which by accurate physiological
investigation may be distinguished in the unicellular Radiolarian organism,
may be distributed in the above indicated conventional fashion into a few
larger and several smaller groups ; it must always be borne in mind,
however, that these overlap in many respects, and that the division of
labour among the different organs in these Protista is somewhat complicated,
notwithstanding
the apparent simplicity of their unicellular organisation. A general classification
of the groups of functions is difficult, because each individual organ
discharges several different functions. Thus the central capsule is pre-eminently
the organ of reproduction and inheritance, but not less (though less conspicuous)
is its importance as the psychical central organ, the unit regulating the
processes of sensation, motion, and also nutrition. In this last respect
it is comparable to the nerve-centres of the Metazoa, whilst the peripheral
nervous system of the latter (including the organs of sense and the muscles)
are in the present instance represented by the pseudopodia, which are at
the same time the most important organs of nutrition and adaptation. In
the calymma also in similar fashion several different physiological functions
are united.
203. Metastasis.-The functions of metastasis and nutrition have in all Radiolaria a purely animal character, so that these Rhizopoda from the physiological standpoint are to be regarded as truly unicellular animals, or Protozoa ("Urthiere "). Since they do not possess, like plants, the power of forming synthetically the compounds (protoplasm, carbohydrates, &c.) necessary for their sustenance, they are compelled to obtain them ready-formed from other organisms. Like other true animals they evolve carbon dioxide by the partial oxidation of those products, and hence they successively take up the oxygen necessary to their existence from their environment.
The question whether the Radiolaria are to be regarded as true animals I discussed fully from various points of view in 1862, and finally answered in the affirmative (L. N. 16, pp. 159-165). Afterwards, when in my Generelle Morphologie (1866) I sought to establish the kingdom Protista, I removed the Radiolaria along with the other Rhizopoda from the animal kingdom proper and placed them in the kingdom Protista (Bd. i. pp. 215-220 ; Bd. ii. p. xxix). Compare also my Protistenreich (L N. 32) and my Natürliche Schöpfungsgeschichte (vii. Aufl., 1879, p. 364). Both these steps appear fully justified when considered in the light of our present increased knowledge. From the physiological standpoint the Radiolaria appear as unicellular animals, for in this respect the animal character of their metastasis (that proper to an oxidising organism) furnishes the sole criterion. On the other hand, from the morphological standpoint, they are to be classed as neutral Protista, for in this respect their unicellular character is the prominent feature, and distinguishes them from all true multicellular animals (Metazoa). Compare my Gastraea Theorie (1873, Jena. Zeitschr. f¨ür Naturwiss., Bd. viii. pp. 29, 53).
204. Nutrition.-The nutritive materials which the Radiolaria require for their sustenance, especially albuminates (plasma) and carbohydrates (starch, &c.), they obtain partly from foreign organisms which they capture and digest, and partly directly from the Xanthellae or Philozoa, the unicellular Algae, with which they live in symbiosis (§ 205). Zooxanthella intracapsularis, found in the ACANTHARIA (§ 76), is probably of the same significance in this respect as Zooxanthella extracapsularis of the SPUMELLARIA and NASSELLARIA (§ 90) ; and perhaps the same is true also of Phaeodella extracpsularis (or Zoochlorella phaeodaris ?) of the PHAEODARIA (§ 89). The considerable quantity of starch or amyloid bodies, elaborated by these inquiline symbiontes, as well as their protoplasm and nucleus, are available, on their death, for the nutrition of the Radiolaria which harbour them. Nutrition by means of other particles obtained by the pseudopodia from the surrounding medium is by no means excluded ; indeed it may be regarded as certain that numerous Radiolaria (especially such as contain no symbiotic Algoid cells) are nourished for the most part or exclusively by this means. Diatoms, Infusoria, Thalamophora (Foraminifera) as well as decaying particles of animal and vegetable tissues can be seized directly by the pseudopodia and conveyed either to the sarcodictyum (on the surface of the calymma) or to the sarcomatrix (on the surface of the central capsule) in order to undergo digestion there. The indigestible constituents (siliceous shells of Diatoms and Tintinnoidea, calcareous shells of small Monothalamia and Polythalamia, &c.) are here collected often in large numbers and removed by the streaming of the protoplasm.
The inception and digestion of nutriment, as it usually appears to take
place by the pseudopodia, has already been so fully treated in my Monograph
(L. N. 16, pp. 135-140), and since then in my paper on the sarcode body
of the Rhizopoda (L. N. 19, p. 342), that I have nothing of importance
to add. Quite recently Karl Brandt has expressed a doubt as to whether
the taking up of formed particles by the pseudopodia and their aggregation
in the calymma be really connected with the process of nutrition. He is
disposed rather to believe that these foreign bodies are usually only accidentally
and mechanically brought into the calymma, and that the nourishment of
the Radiolaria is derived exclusively or pre-eminently from the symbiotic
Xanthellae (L. N. 52, pp. 88-93). I must, however, maintain my former opinion,
which I have only modified insomuch that I now regard the sarcodictyum
(on the outer surface of the calymma, § 94) rather than the sarcomatrix
(on the outer surface of the central capsule, § 92) as the principal
seat of true digestion and assimilation. From the sarcodictyum the dissolved
and assimilated nutritive matters. may pass by the intracalymmar pseudopodia
(or sarcoplegma, § 93) into the sarcomatrix, and hence may reach the
endoplasm through the openings in the central capsule. To what an extent
the Radiolaria are capable of taking up even large formed bodies into the
calymma, is shown by the striking instance of Thalassicolla sanguinolenta,
which
becomes so deformed by the inception of numerous coccospheres and coccoliths,
that I described it as a special genus under the name
Myxobrachia (compare
pp. 23, 30 ; also L. N. 21, p. 519, Taf. xviii., and L. N. 33, p.
37).
205. Symbiosis.-Very many Radiolaria, but by no means all members of this class, live in a definite commensal relation with yellow unicellular Algae of the group Xanthellae. In the ACANTHARIA they live within the central capsule (Zooxanthella intracapsularis, § 76), in the SPUMELLARIA and NASSELLARIA, on the other hand, within the calymma but outside the central capsule (Zooxanthella extracapsularis, § 90) ; in the PHAEODARIA a special form of these symbiotic unicellular Algae appears to inhabit the phaeodium in the extracapsulum, and to compose a considerable portion of the phaeodellae (Zooxanthella phaeodaris, § 90, or better perhaps Zoochlorella phaeodaris, § 89). Undoubtedly this commensal life is in very many cases of the greatest physiological significance for both the symbiontes, for the animal Radiolarian cells furnish the inquiline Xanthellae not only with shelter and protection, but also with carbon dioxide and other products of decomposition for their nutriment ; whilst on the other hand the vegetable cells of the Xanthellae yield the Radiolarian host its most im portant supply of nutriment, protoplasm and starch, as well as oxygen for respiration. Hence it is not only theoretically possible, but has been experimentally proved, that Radiolaria which contain numerous Xanthellae can exist without extraneous nutriment for a long period in closed vessels of filtered sea-water, kept exposed to the sunlight ; the two symbiontes furnish each other mutually with nourishment, and are physiologically supplementary to each other by reason of the opposite nature of their metastasis. This symbiosis is not necessary, however, for the existence of the Radiolaria ; for in many species the number of Xanthellae is very variable and in many others they are entirely wanting.
The symbiosis of the Radiolaria and Xanthelke, or "yellow cells" (§§ 76, 90) was first discovered by Cienkowski in 1871 (L. N. 22). Ten years later this important and often doubted fact was established by extended observations and experiments almost simultaneously by Karl Brandt (L. N. 38, 39) and Patrick Geddes (L. N. 42, 43). This commensal life may be compared with that of the lichens, in which an organism with vegetable metastasis (the Algoid gonidia) and an organism with animal metastasis (the Fungoid hyphae) are intimately united for mutual benefit. But the symbiosis of the Xanthellae and Radiolaria is not as in the lichens a phenomenon essential for their development, but has more or less the character of an accidental association. The number of the inquiline Xanthellae is so variable even in one and the same species of Radiolaria, that they do not appear to be exactly essential to its welfare ; and in many species they are entirely wanting. Their significance is questionable in the case of those numerous deep-sea Radiolaria which live in complete darkness, and in which, therefore, the Xanthelke, even if present, could excrete no oxygen on account of the want of light. Nevertheless it is possible that the phaeodellae of the PHAEODARIA (usually green, olive, or brown in colour), which are true cells, represent vegetable symbiontes, which in the absence of sunlight are able to evolve oxygen by the aid of the phosphoresence of other abyssal animals. Since the PHAEODARIA are, for the most part, dwellers in the deep-sea, and since the voluminous phaeodium must be of great physiological importance, a positive solution of this hypothetical question would be of no small interest (compare § 89).
206. Respiration.-The respiration of the Radiolaria is animal in nature, since all Protista of this class, like all other true Rhizopoda, take in oxygen and give off carbon dioxide. Probably this process goes on continuously and is tolerably active, as may be inferred from the fact that Radiolaria cannot be kept for long in small vessels of sea water unless either they contain numerous Xanthellae or the water is well aërated. The oxygen is obtained from two sources, either from the surrounding water or from the enclosed Xanthellae, which in sunlight evolve considerable quantities of this gas. Correspondingly, the carbon dioxide which is formed during the process of oxidation of the Radiolaria is either given up to the surrounding water or to the inquiline Xanthellae, which utilise it for their own sustenance (§§ 204, 205).
The significance of the symbiotic Xanthellae for the respiration of
the enclosing Radiolaria may be shown experimentally in the following way.
If two Polycyttarian colonies of equal size, both of which contain numerous
Xanthellae, be placed in equal quantities of filtered sea-water in sealed
glass tubes, and if one tube be placed in the dark the other in the light,
the colony in the former rapidly perishes, but not that in the latter ;
the
Xanthellae excrete only under the influence of sunlight the oxygen necessary
for the life of the Radiolarian (compare Patrick Geddes, L. N. 42, p. 304).
207. Circulation.-In the protoplasm of all Radiolaria, both inside and outside the central capsule, slow currents may be recognised which fall under the general term circulation, and have already been compared to the cycbosis in the interior of animal and vegetable cells, as well as to the sarcode streams in the body of other Rhizopoda. These plasmatic currents or "plasmorrheumata" probably continue throughout the whole life of the Radiolaria, and are of fundamental importance for the performance of their vital functions. They depend upon slow displacements of the molecules of the plasma (plastidules or micellae) and cause a uniform distribution of the absorbed nutriment and a certain equalisation of the metastasis. Furthermore they are of great importance also in the inception of nutriment, the formation of the skeleton, locomotion, &c. Sometimes the circulation is directly perceptible in the plasma itself ; but usually it is only visible owing to the presence of granules (sarcogranula), which are suspended in the plasma in larger or smaller numbers. The movements of these granules are usually regarded as passive, due to the active displacement of the molecules of the plasma. Although the intracapsular protoplasm is in communication with the extracapsular through the openings in the capsule membrane, nevertheless the currents exhibit certain differences in the two portions of the malacoma. It is sometimes possible, however, to recognise the direct connection between them and to observe how the granules pass through the openings in the capsule-membrane.
208. Currents in the Endoplasm.- Intracapsular circulation or
a certain slow flowing of the plasma within the central capsule is probably
just as common in the Radiolaria as without it, but it is not so easy to
observe in the former case as in the latter. A more satisfactory proof
of these endoplasmatic currents is furnished by the arrangement of the
protoplasm within the central capsule, since this is (at all events in
part) an effect produced by them. In this respect the two main divisions
of the class show characteristic differences. In the Porubosa (the SPUMELLARIA,
§ 77, and the ACANTHARIA, § 78) the endoplasm is in general distinguished
by a more or less distinct radial structure, which is to be regarded as
the effect of alternating centripetal and centrifugal radial streams. In
the Osculosa, on the other hand, this radial structure is absent and the
intracapsular plasmatic streams converge or diverge towards the osculum
or main opening in the central capsule which lies at the basal pole of
its main axis, and through which the mass of the endoplasm issues into
the calymma. The two legions of the Osculosa, however, present differences
in this respect. In the NASSELLARIA (§ 79) the endoplasmatic currents
appear to unite in an axial main stream at the apex of the monaxon central
capsule, and this apical stream seems to split into a conical bundle, the
individual threads of which pass diverging between the myophane fibrillae
of the podoconus towards the basis of the central capsule, and issue through
the pores of the porochora. In the PHAEODARIA (§ 80), on the other
hand, meridional currents of endoplasm are probably present on the inner
surface of the capsule, which flow from the aboral pole of the vertical
main axis to its basal pole, and return in the reverse direction.
209. Currents in the Expolasm.-Extracapsular circulation, or a distinct flowing of the plasma outside the central capsule, may be readily observed in all Radiolaria which are examined alive ; this is most readily seen in the astropodia, or those free pseudopodia which radiate from the sarcodictyum on the surface of the calymma into the surroundinig water. The granular movement is often quite as clear in the sarcodictyum itself, and may be recognised in the collopodia, which compose the irregular plasmatic network within the calymma. More rarely it is possible to follow the granular stream thence through the sarcomatrix, and further into the interior of the central capsule. In general the direction of the extracapsular protoplasmic streams is radial, and it is frequently possible, even in a single free astropodium, to observe two streams opposite in direction, the granules on one side of the radial sarcode thread moving centripetally, those on the other side centrifugally. If the threads branch, and neighbouring ones become united by connecting threads, the circulation of the granules may proceed quite irregularly in the network thus formed. The rapidity and character of the extracapsular currents are subject to great variations.
The different forms of extracapsular sarcode currents have been already very fully described in my Monograph (L. N. 16, pp. 89-126), and in my critical essay on the sarcode body of the Rhizopoda (L. N. 19, p. 357, Taf. XXVI.).
210. Secretion.-Under the name secretions, in the strict sense, all the skeletal formations of the Radiolaria may be included. They may be divided according to their chemical composition into three different groups: pure silica in the SPUMELLARIA and NASSELLARIA, a silicate of carbon in the PHAEODARIA, and acanthin in the ACANTHARIA (compare § 102). It may indeed be assumed that these skeletons arise directly by a chemical metamorphosis (silicification, acanthinosis, &c.) of the pseudopodia and protoplasmic network ; and this view seems especially justified in the case of the Astroid skeleton of the ACANTHARIA (§ 114), the Spongoid skeleton of the SPUMELLARIA (§ 126), the Plectoid skeleton of the NASSELLARIA (§ 125), the Cannoid skeleton of the PHAEODARIA (§ 127), and several other types. On closer investigation, however, it appears yet more probable that the skeleton does not arise by direct chemical metamorphosis of the protoplasm, but by secretion from it ; for when the dissolved skeletal material (silica, acanthin) passes from the fluid into the solid state, it does not appear as imbedded in the plasma, but as deposited from it. However, it must be borne in mind that a hard line of demarcation can scarcely, if at all, be drawn between these two processes. In the ACANTHARIA the intracapsular sarcode is the original organ of secretion of the skeleton ; in the other three legions, on the other hand, the extracapsulum performs this function (§§ 106, 107). In addition to the skeleton, we may regard as secretions (or excretions) the intracapsular crystals (§ 75) and concretions (§ 75A), and perhaps certain pigment-bodies (§§ 74, 88) ; and further, the calymma (§ 82) may be considered to be a gelatinous secretion of the central capsule, and perhaps also the capsule-membrane, in so far as it represents only a secondary excretory product of the unicellular organism.
211. Adaptation.-The innumerable and very various adaptive phenomena which we meet with in the morphology of the Radiolaria, and especially in that of their skeleton, are like other phenomena of the same kind, to be ultimately referred to altered nutritional relations. These may be caused directly either by the influence of external conditions of existence (nutrition, light, temperature, &c.), or by the proper activity of the unicellular organism (use or disuse of its organs, &c.), or, finally, by the combined action of both causes in the struggle for existence. In very many cases the cause to which the origin of a particular form of Radiolaria is due may be directly perceived or at least guessed at with considerable probability ; thus, for example, the lattice-shells may be explained as protective coverings, the radial spines as defensive weapons, and the anchor-hooks and spathillae as organs of prehension, which are of advantage to their possessors in the struggle for existence ; the regular arrangement of the radial spines in the Radiolaria may also be explained on hydrostatic grounds, it being advantageous that the body should be maintained in a definite position of equilibrium, &c. The well-known laws of direct or actual adaptation, which we designate cumulative, correlative, divergent adaptation, &c., here explain a multitude of morphological phenomena. The connection is less distinct in the case of the laws of indirect or potential adaptation, although this must play as important a part in the formation of the Radiolaria as in that of other organisms (compare on this head my Generelle Morphobogie, Bd. ii. pp. 202-222).
212. Reproduction.-The most common form of reproduction in the Radiolaria is the formation of spores in the central capsule, which in this respect is to be regarded as a sporangium (§ 215). In many Radiolaria (Polycyttaria and PHAEODARIA), however, there occurs in addition an increase of the unicellular organism by simple division (§ 213); upon this the formation of colonies in the social Radiolaria is dependent (§ 14). Reproduction by gemmation is much less common, and has hitherto been observed only in the Polycyttaria (§ 214). In this group alone there also occur at certain times two different forms of swarm-spores which copulate, and thus indicate the commencement of sexual reproduction (Alternation of Generations, § 216). The general organ of reproduction is in all cases the central capsule, whilst the extracapsulum never takes an active part in the process.
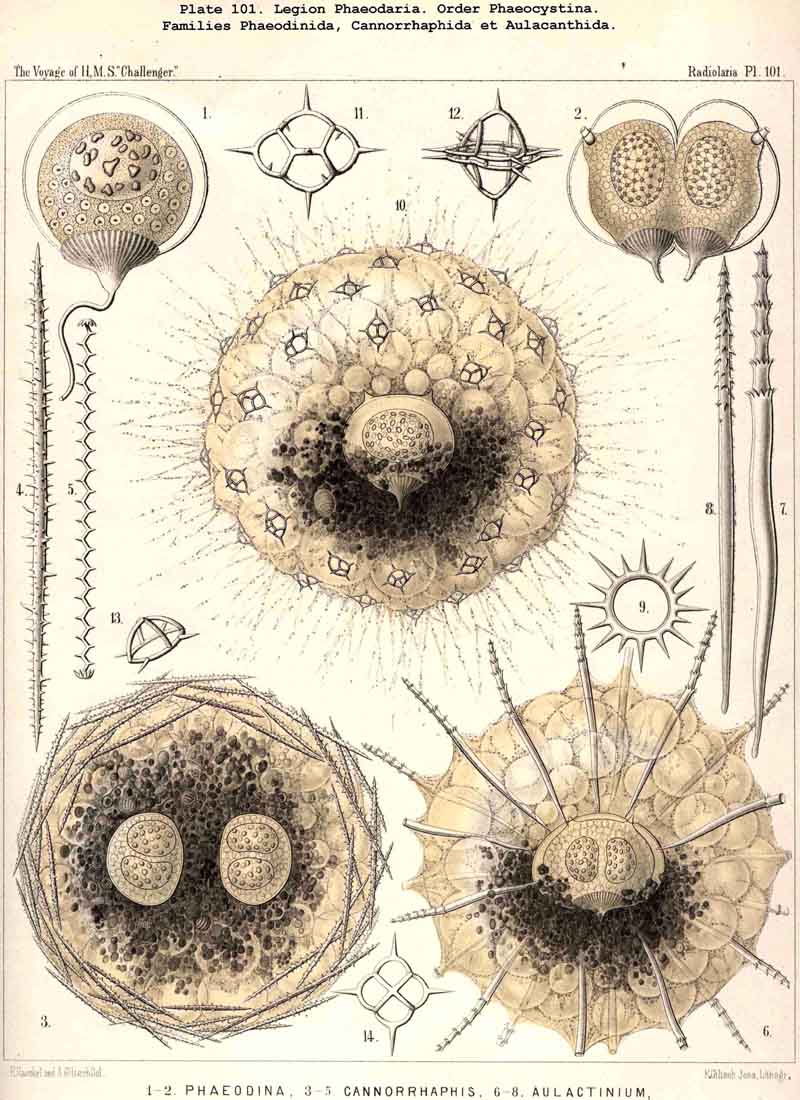
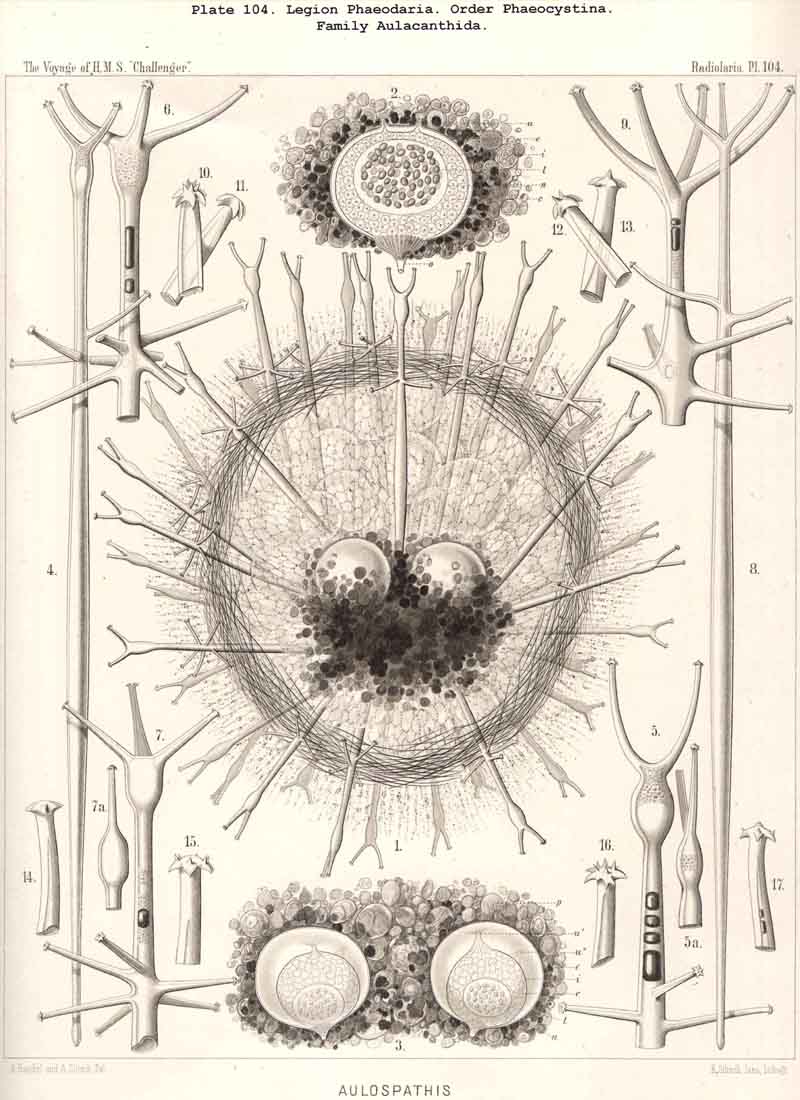
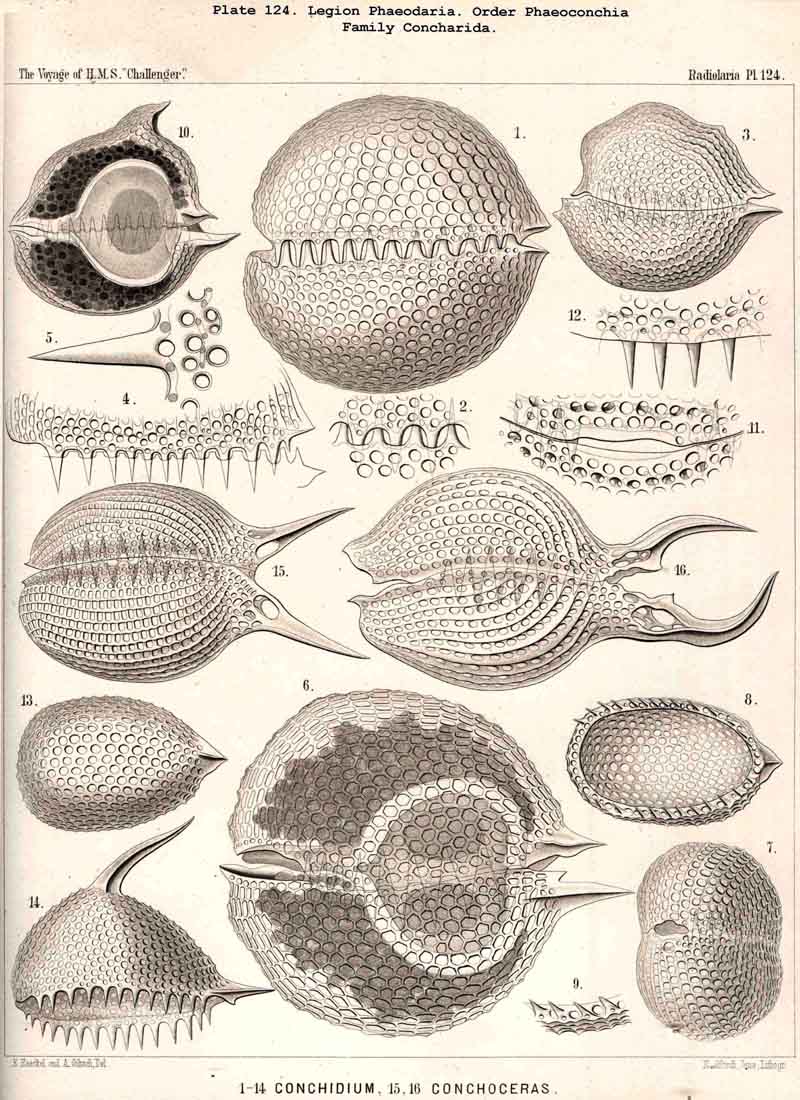
213. Cell-Division.-Increase by cell-division among the Radiolaria in the early stage, before the formation of the skeleton, is widely distributed (perhaps even general ?) ; in the adults of this class it is rather rare and limited to certain groups. It is most readily observed in the Polycyttaria ; the growth of the colonies in this social group depends mainly (and in many species exclusively) upon repeated spontaneous division of the central capsule ; all the individuals of each colony (in so far as this has not arisen by the accidental fusion of two or more colonies) are descendants of a single central capsule, which has arisen from an asexual swarm-spore (§ 215) or from the copulation of two sexual swarm-spores (§ 216). Whilst the central capsules of the colonies continually increase by division, their calymma remains a common gelatinous sheath. Among the SPUMELLARIA reproduction by simple cell-division probably occurs also in many monozootic Collodaria. Among the ACANTHARIA the peculiar group Litholophida has perhaps arisen by the spontaneous division of Acanthonida (see p. 734). Among the PHAEODARIA increase by cell-division seems to occur commonly in many groups, as in the Phaeocystina, which have no skeleton (Phaeodinida, Pl. 101, fig. 2), or only an incomplete Beloid skeleton (Cannorrhaphida, Pl. 101, figs. 3, 6, and Aulacanthida, Pl. 104, figs. 1-3). The Phaeosphaeria also (Aulosphaerida, Coelacanthida) and the Phaeogromia (Tuscarorida, Challengerida) appear sometimes to divide ; at all events, their central capsule often contains two nuclei. Of special interest is the spontaneous division of the Phaeoconchia, especially the Concharida (Pl. 124, fig. 6). In all monozootic Radiolaria, the nucleus first divides by a median constriction into two equal halves (usually by the mode of direct division) ; then the central capsule becomes constricted in the middle (in the PHAEODARIA in the vertical main axis), and each portion of the capsule retains its own nucleus. In the Phaeoconchia each half or daughter-cell corresponds to one valve of the shell, dorsal or ventral, so that probably on subsequent separation each daughter-cell retains one valve of the mother-cell, and forms a new one for itself by regeneration (as in the Diatoms). In the polyzootic Radiolaria, which already contain many small nuclei, but usually only a single central oil-globule in each central capsule, the division of the latter is preceded by that of the oil-globule. In many Polycyttaria the colony as a whole multiplies by division.
The increase of the central capsule by division was first described in 1862 in my Monograph (L. N. 16, p. 146) ; since then R. Hertwig (L. N. 26, p. 24) and K. Brandt (L. N. 52, p. 144) have confirmed my statement. In the PHAEODARIA the division of the central capsule appears always to take place in the main axis ; in the bilateral sometimes in the sagittal, sometimes in the frontal plane. In the Tripylea each daughter-cell seems to retain one parapyle and half the astropyle (compare the general description of the PHAEODARIA, Pl. 101, figs. 1-6, Pl. 104, figs. 1-3, and also Hertwig, L. N. 33, p. 100, Taf. x. figs. 2, 11). Regarding the spontaneous division of colonies of the Polycyttaria, see K. Brandt, L. N. 52, p. 142.
214. Cell-Gemmation.-Reproduction by gemmation has hitherto been observed only in the social Radiolaria, but in them it appears to be widely distributed, and in very young colonies is perhaps almost universally present. The gemmules or capsular buds (hitherto described as "extracapsular bodies") are developed on the surface of young central capsules before these had secreted a membrane. They grow usually in considerable numbers, from the surface of the central capsule, which is some times quite covered with them. Each bud usually contains a raspberry-like bunch of shining fatty globules, and by means of reagents a few larger or a considerable number of smaller nuclei may be recognised in them ; the naked protoplasmic body of the bud is not enclosed by any membrane. As soon as the buds have reached a certain size they are constricted off from the central capsule and separated from it, being distributed in the meshes of the sarcoplegma by the currents in the exoplasm. Afterwards each bud becomes developed into a complete central capsule by surrounding itself with a membrane when it has attained a definite size. From the special relations of the processes of nuclear formation, which take place in the multiplication of the social central capsules by gemmation and by cell-division, it would appear that the capsules produced by the former method afterwards produce anisospores, whilst those in the latter way yield isospores (§ 216).
The gemmules or capsular buds of the Polycyttaria were first accurately described by Richard Hertwig (L. N. 26, pp. 37-39), under the name "extracapsular bodies," and their significance rightly indicated ; earlier observers had incidentally mentioned and figured them, but had not seen their origin from the central capsule. Quite recently Karl Brandt has given a very painstaking account of them in the different Polycyttarian genera (L. N. 52, pp. 179-198). In the Monocyttaria such a formation of buds has not yet been observed. The basal lobes of the central capsule, which occur in many NASSELLARIA, are not buds, but simple processes of the capsule, due to its protrusion through the collar pores of the cortinar septum (§ 55).
215. Sporification.-Asexual reproduction by the formation of movable flagellate spores has been hitherto observed only in a very small number of genera ; but since these belong to very different groups, and since the comparative morphology of the capsule appears to be similar throughout as regards the structure and development of its contents, it may be safely assumed that this kind of reproduction occurs quite generally in the Radiolaria. In all cases it is the contents of the central capsule, from which the swarm-spores are formed, both nucleus and endoplasm taking an equal share in the process ; in all cases the spores produced are very numerous, small, ovoid or reniform, and have one or two very long slender flagella at one extremity (see §§ 141, 142). Since the whole contents of the mature central capsule are used up in the formation of these flagellate zoospores, it discharges the function of a sporangium. The division of the simple primary nucleus into numerous small nuclei, which usually (serotinous Radiolaria) takes place only shortly before sporification, but sometimes (precocious Radiolaria, § 63) happens very early; is the commencement of the often repeated process of nuclear division, which terminates with the production of a very large number of small spore-nuclei. The nucleolus often divides very peculiarly (§ 69, C). Each spore nucleus becomes surrounded by a portion of endoplasm and usually receives in addition one or more fatty granules, and sometimes also a small crystal (hence the "crystal-spores "). The size of the flagellate zoospores which emerge from the ruptured central capsule and swim freely in the water by means of their flagellum, varies generally between 0.004 and 0.008 mm. The extracapsubum is not directly concerned in the sporification, but undergoes degeneration during the process and perishes at its conclusion.
The first complete and detailed observations on the formation of spores
in the Radiolaria were published by Cienkowski in 1871 and related to two
genera of Polycyttaria, the skeletonless Collozoumn and the spherical-shelled
Collosphaera
(L.
N 22, p. 372, Taf. xxix.), These were subsequently continued and supplemented
by R Hertwig (1876, L. N. 26, pp. 26-42, and L. N. 33 p. 129), and a general
summary of these results has been given by Bütschli (L. N. 41, pp.
449-455). Recently Karl Brandt has given a very detailed and fully illustrated
account of the sporification of the Polycyttaria (L. N. 52, pp. 145-178).
I have also had the opportunity during my sojourn in the Canary Islands
(1866), in the Mediterranean at Corfu (1877), and Portofino (1880), as
well as in Ceylon (1881), of observing the development of flagellate zoospores
from the central capsule of individuals of all four legions: among the
SPUMELLARIA in certain Colloidea, Beloidea, Sphaeroidea, and Discoidea,
among the ACANTHARIA in several Acanthometra and Acanthophracta, among
the NASSELLARIA in individuals belonging to the Stephoidea, Plectoidea,
and Cyrtoidea, and among the PHAEODARIA in one Castanellid. In most zoospores
I could distinctly observe only a single long flagellum ; sometimes,
however, two or even three appeared to be present, but the determination
of their number is very difficult.
216. Alternation of Generations.-A peculiar form of reproduction, which may be designated "alternation of generations," appears to occur generally in the Polycyttaria, but has not yet been observed in the Monocyttaria. All Collozoida, Sphaerozoida, and Collosphaerida which have hitherto been carefully and completely examined with respect to their development, are distinguished by the production of two different kinds of swarm-spores, isospores and anisospores. The Isospores (or monogonous spores) correspond to the ordinary asexual zoospores of the Monocyttaria (§ 215); they possess a homogeneous, doubly refracting nucleus of uniform constitution and develop asexually, without copulation. The Anisospores (or amphigonous spores), on the other hand, are sexually differentiated and possess a heterogeneous, singly refracting nucleus of twofold constitution ; they may therefore be distinguished as female macrospores and male microspores. The Macrospores (or gynospores, comparable with the female macrogonidia of many Cryptogams) are larger, less numerous, and possess larger nuclei, which are less easily stained, and have a fine filiform trabecular network. On the other hand the Microspores (or androspores, comparable with the male microgonidia) are much smaller and more numerous, and are distinguished by their smaller nuclei, which have thicker tuberculae and become stained more intensely. The gynospores and androspores are developed in the Collozoida and Sphaerozoida in the same individual, but not in the Collosphaerida. It is very probable that these two forms of anisospores copulate with each other after their exit from the central capsule and thus produce a new cell by the simplest mode of sexual reproduction. But, since the same Polycyttaria, which produce these anisospores, at other times give rise to ordinary or asexual isospores, it is further possible that these two forms of reproduction alternate with each other, and that the Polycyttaria thus pass through a true alternation of generations. This has not yet been observed in the Monocyttaria, and hence these latter seem to bear to the Polycyttaria a relation similar to that in which the sexless solitary Flagellata (Astasiea) stand to the sexual social Flagellata (Volvocinea). In the two analogous cases the sexual differentiation may be regarded as a consequence of the social life in the gelatinous colonies.
The sexual differentiation of the Polycyttaria was first discovered in 1875 by R. Hertwig, and accurately described in the case of Collozoumm inerme as occurring in addition to the formation of the ordinary crystal-spores (L. N. 26, p. 36) ; compare also the general discussion of Bütschli (L. N. 41, p. 52). Recently Karl Brandt has demonstrated the formation of both homogeneous isospores (crystal-spores) and heterogeneous anisospores (macro- and microspores) in seven different species of Polycyttaria, and has come to the conclusion that in all social Radiolaria there is a regular alternation between the former and latter generations. Compare his elaborate account of the colonial Radiolaria of the Gulf of Naples (L. N. 52, pp. 145-178).
217. Inheritance.-Inheritance is to be regarded as the most important accompaniment to the function of reproduction, and especially in the present case, because the comparative morphology of the Radiolaria furnishes abundant instances of the action of its different laws. The laws of conservative inheritance are illustrated by the comparative anatomy of the larger groups ; thus, in the four legions the characteristic peculiarities of the central capsule are maintained unaltered in consequence of continuous inheritance, although great varieties appear in the skeleton in each legion. The individual parts of the skeleton furnish by their development on the one hand and their degeneration on the other, especially in the smaller groups, examples of progressive inheritance. Thus in the SPUMELLARIA the constant formation of the primary lattice-shell (a central medullary shell) and its ontogenetic relation to the secondary one, which is developed concentricalby round it, can only be explained phylogenetically by conservative inheritance, whilst on the other hand the characteristic differentiation of the axes in the various families of the SPUMELLARIA is to be explained by progressive inheritance. In the ACANTHARIA the arrangement of the twenty radial spines (in accordance with Müller's law, §§ 110, 172) was first acquired by a group of the most archaic Actinelida (Adelacantha) through hydrostatic adaptation, and has since been transmitted by inheritance to all the other families of the legion (Icosacantha). The morphology of the NASSELLARIA is not less interesting, because here several different heritable elements (the primary sagittal ring and the basal tripod) combine in the most manifold ways in the formation of the skeleton (compare §§ 123, 124, 182). The affinities of the genera in the different families yield an astonishing variety of interesting morphological phenomena, which can only be explained by progressive inheritance. The same is true also of the PHAEODARIA. In this legion the primary inheritance is especially manifested in the constant and firm structure of the central capsule with its characteristic double wall and astropyle, whilst the formation of the skeleton in this legion proceeds in different directions by means of divergent adaptation. The morphology of the Radiolaria thus proves itself a rich source of materials for the physiological study of adaptation and inheritance.
{kind=link}
{kind=link}
{kind=link}