 |
(§§ 153-200.)
153. Sources of Phylogenetic Knowledge.-For the purpose
of constructing a hypothetical genealogical tree of the Radiolaria, as
of all other organisms, three sources of information are open to us, viz.,
palaeontology, comparative ontogeny, and comparative anatomy. In the present
case, however, these three sources are of very different value ; the
first two are at present only very inadequately known and have only been
partially investigated, hence they can only be utilised to a very slight
extent. The comparative anatomy of the Radiolaria, on the other hand, is
so completely known, and affords such certain glimpses into the morphological
relations of the related groups, that by its aid we are in a position at
all events to lay down the general features of their phylogeny with some
probability, and to lay the foundation of a natural system.
154. Natural and Artificial Systems.-Although in the classification of the Radiolaria, as in the case of all other organisms, the natural system must be regarded as the goal of systematic classification, our phylogenetic knowledge of the Radiolaria is too fragmentary and inadequate to admit of the systematic arrangement here adopted being regarded as a thoroughly consistent natural system, that is, as representing the true genealogical tree of the class. Owing, however, to the extraordinary variety of form of the Radiolaria, and the complicated relationships of the larger and smaller groups, a synoptical grouping of the different categories and the erection of a complete, even if to some extent artificial, system, becomes a logical necessity. Under these circumstances, and regard being had to both these conditions, the following systematic treatment of the Radiolaria will appear as a compromise between the natural and artificial systems, like all other zoological and botanical classificatory attempts. On the one hand, the attempt is made to arrange the larger and smaller groups as nearby as possible according to their phylogenetic relationships, whilst, on the other hand, the practice of circumscribing each by a definition as clear and logical as possible has been carried out. Since these two efforts naturally often come into contact, the insufficiency of many parts of the arrangement is obvious, hence its hypothetical and provisional character is emphatically stated.
155. Systematic Categories.-The categories or different
orders of divisions have in the Radiolaria, as in all other organisms,
no absolute
significance, but only a relative value. In itself
it is quite unimportant whether the whole group be regarded, as at first,
as a family
(Ehrenberg, 1847), or as an order (J. Müller,
1858), or as a class (Haeckel, 1881). These different views are
regulated, on the one hand, by the known extent of the group and by the
amount of our acquaintance with it, and on the other, by comparison with
related groups and by reference to their conventional disposition. When,
therefore, the whole class, Radiolaria, is here divided into two subclasses,
four legions, eight orders, eighty-five families, &c., these artificial
categories are drawn up only in the conviction that by this means the easiest
survey and most thorough insight into the system as a whole may be attained
; this latter will indeed approach as far as possible the ideal of a natural
system, but must on numerous practical grounds always remain more or less
artificial. Since it is to be expected that with the progress of our systematic
knowledge the rank of the various categories will rise, it is possible
that in the future the arrangement of the group may be somewhat as follows
:-Phylum,
RADIOLARIA
; Four Classes, SPUMELLARIA, NASSELLARIA, ACANTHARIA, PHAEODARIA
; Eight Legions (Nos. 1.-VIII. in the following Table);
Twenty
Orders (Nos. 1-20 in the Table), &c.
|
|
 |
156. Formation of Species.-The totality of similar forms, which we unite in one species, and which in the earlier dogmatic systems was regarded as a category of absolute value, possesses only a relative value like all other systematic categories (§ 155). According to the individual views of the systematist and the general survey which he has attained of the smaller and larger systematic groups, the conception of a species adopted in his practical work will be wider or narrower. In the present systematic arrangement a medium extent has been adopted. It is shown that in the Radiolaria, as in all other extensive groups of organisms, the constancy of the species is very variable in the different groups. Many families of Radiolaria are very rich in "bad species," i.e., very variable forms, in which the process of the formation of species is seen in progress ; such, for example, areamong the SPUMELLARIA, the Sphaerozoida, Stylosphaerida, Phacodiscida and Pylonida ; among the ACANTHARIA, the Amphilonchida and Phractopeltida ; among the NASSELLARIA, the Stephoidea and Botryodea; and among the PHAEODARIA, the Aulacanthida, Sagosphaerida, Castanellida and Concharida. On the other hand, in some families numerous "good species" may be distinguishied, since the intermediate connecting forms are no longer present and the forms have become relatively constant. As instances of such families may be mentioned, among the SPUMELLARIA, the Astrosphaerida, Cyphinida, Porodiscida and Tholonida ; among the ACANTHARIA the Quadrilonchida and Dorataspida ; among the NASSELLARIA, the Spyroidea and Cyrtoidea ; among the PHAEODARIA, the Challengerida, Medusettida, Circoporida and Coelographida. The more carefully the different groups are studied, the more numerous the individuals of each species under comparison, the greater becomes the number of "bad" species among the Radiolaria, and the smaller, the number of good ones. Originally, no doubt, all "species bonae" were "malae." There may be observed in the manifold skeletal forms of the Radiolaria, on the one hand, the utmost accuracy of con figuration, and on the other, the greatest variability, and hence a careful comparative study of them leads to a firm conviction of the gradual "Transformation of Species," and of the truth of the "Theory of Descent."
157. Palaeontological Development.-The palaeontology of the Radiolaria already offers very considerable material for study ; but in consequence of its incompleteness this is of little value for the study of the phylogeny of the class. By far the larger portion of the fossil Radiolaria belong to the Tertiary period ; only quite recently have numerous well-preserved fossil Radiolaria been described from the Mesozoic period, and especially from the Jura. Of Palaeozoic Radiolaria (from the coal measures) only slight traces are known. Moreover, the fossil Radiolaria hitherto known have been found only in very circumscribed and widely separated localities. The majority of all the species belong to the small island of Barbados. Although our palaeontobogical acquaintance with the Radiolaria must necessarily be incomplete for this reason, it is still more so since at least thirty out of the eighty-five families (that is more than a third) could not possibly leave any fossil remains, either because they possess no skeleton, or because of its chemical composition.
Of the four legions of the Radiolaria, the ACANTHARIA (on account of the solubility of their astroid acanthin skeletons) have entirely vanished and have never been found fossil. Of the PHAEODARIA, whose silicate skeleton is not as a rule capable of fossilisation, only one section (Dictyochida) of a single family (Cannorrhaphida) has been observed fossil. Hence the fossil remains of the Radiolaria belong almost exclusively to the two legions, SPUMELLARIA and NASSELLARIA, which were formerly united under the term "Polycystina." Among these, however, the skeletonless Thalassicollida, Collozoida, and Nassellida could leave no traces. Hence there only remain fifty-five families of which we might expect to find fossil siliceous skeletons. Even of these, however, scarcely the half are certainly known in the fossil condition, whilst of the remainder nothing certain is known ; for example, of the large order Larcoidea (among the SPUMELLARIA) and of the Stephoidea (among the NASSELLARIA) with a few isolated exceptions, no fossils are known. The great majority of fossil Radiolaria belong to the two NASSELLARIAN orders Cyrtoidea and Spyroidea (two relatively very highly developed groups); next to these follow the orders Discoidea and sphaeroidea among the SPUMELLARIA. From these palaeontological facts it is obvious that our present very incomplete acquaintance with the fossil Radiolaria is quite insufficient to warrant us in drawing any conclusions from it regarding the phylogenetic development or palaeontological succession of the individual groups.
158. Origin of the Four Legions.-The agreement of all Radiolaria in those constant and essential characters of the unicellular body, which distinguish them from all other Protista (especially the differentiation of the malacoma into a central capsule and extracapsulum), justifies the conclusion that all members of this class have been developed from a common undifferentiated stem-form. Only the simplest form of the SPUMELLARIA, a skeletonless spherical cell with concentric spherical nucleus and calymma, can be regarded as such. The simplest form of the Thalassicollida which is now extant (Actissa, Procyttarium, p. 12), corresponds so exactly to the morphological idea of that hypothetical stem-form that it may unhesitatingly be regarded in a natural system as the common point of origin of the whole class. On the other hand, Actissa is so closely related to the simple Heliozoa (Actinophrys, Actinosphaerium, Heterophrys, Sphaerastrum, &c.) that its origin from this group of Rhizopoda is exceedingly probable. The three legions ACANTHARIA, NASSELLARIA, and PHAEODARIA are to be regarded as three main diverging branches of the genealogical tree, which have been developed in different directions and are only connected by their simplest stem-forms (Actinelius,Nassella, Phaeodina) with the stem-form of the SPUMELLARIA, the primordial Actissa.
159. Phylogeny of the Spumellaria.-The legion SPUMELLARIA or PERIPYLEA is to be regarded as the common stem-group of the Radiolaria, and its simplest form, Actissa, as the primitive genus or radical form of the whole class ; for it possesses in the simplest and most undifferentiated form all those characters by which the Radiolaria are distinguished from other Protista ; all the other genera of the class may be derived from it by succes sive modifications. Considered as a legion the whole group SPUMELLARIA is undoubtedly monophyletic, for all its members possess those essential characters by which it is dis tinctively marked off from the other three legions, more especially a simple capsule-membrane, which is everywhere evenly perforated by innumerable small pores ; the nucleus lies originally in the centre of the spherical central capsule. Furthermore, all SPUMELLARIA lack those positive characters which distinguish the three remaining legions the centrogenous acanthin skeleton of the ACANTHARIA, the basal porochora and the monaxon podoconus of the NASSELLARIA, the astropyle and phaeodium of the PHAEODARIA.
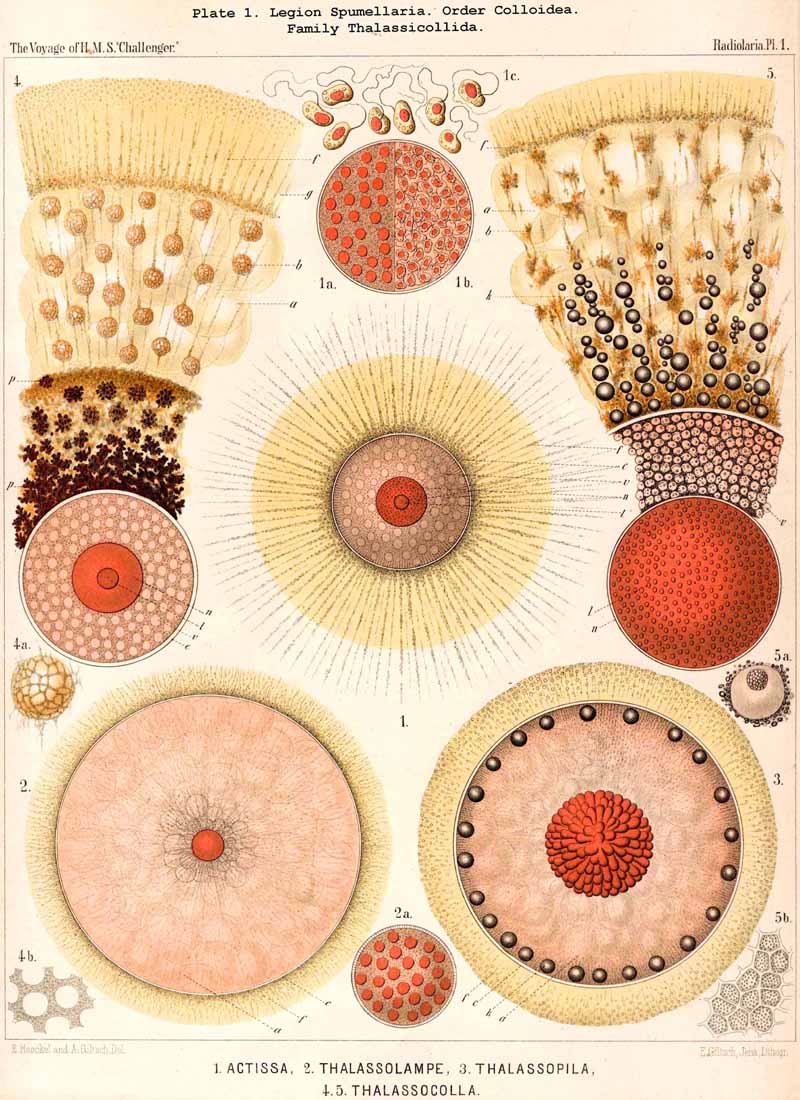
160. Origin of the Spumellaria.-The genus Actissa (p. 12, Pl. 1, fig. 1) presents the Radiolarian type in its simplest and most primitive form a spherical central capsule, which encloses in its middle a spherical nucleus, and which is surrounded by a spherical calymma. The whole unicellular body consists, therefore, of three concentric spheres, and possesses neither skeleton nor alveoles, nor other differentiated parts. The innumerable fine pseudopodia, which issue from the central capsule through the evenly distributed pores in its membrane, radiate in all directions through the calymma and pass out over its surface. Actissa can, therefore, be directly derived phylogenetically from the simplest skeletonless Heliozoa (Actinophrys, Heterophrys, Actinosphaerium, Sphaerastrum). The only essential difference between the two consists in the development of the central capsule, which in Actissa separates as a distinct membrane the endoplasm from the exoplasm. This differentiation, which we regard as the most important distinguishing character of the Radiolaria, has been transmitted by inheritance, along with the formation of flagellate spores in the central capsule, from Actissa, the primitive parent, to all the other Radiolaria.
161. Hypothetical Genealogical Tree of the Spumellaria.
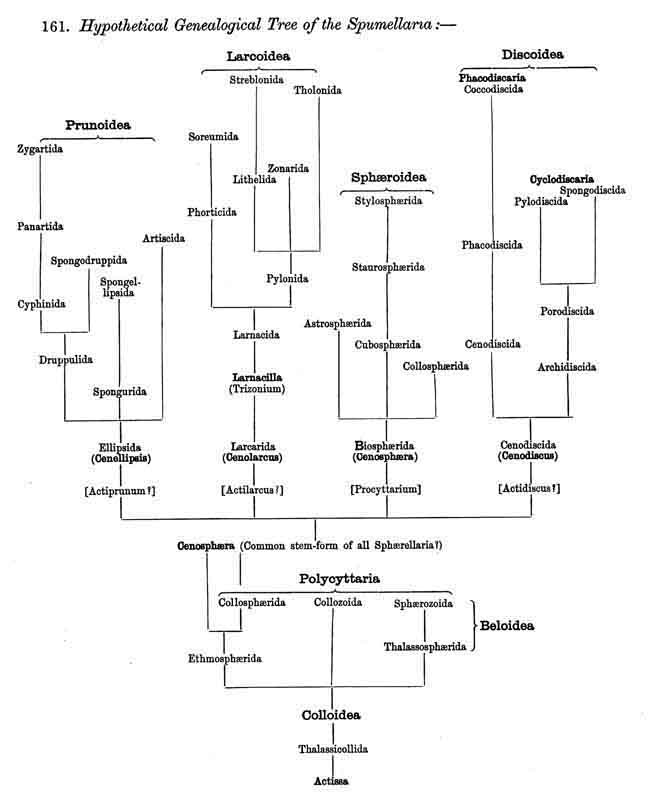 |
162. Collodaria and Sphaerellaria.-Whilst in all SPUMELLARIA the malacoma agrees in possessing the characteristic features of the legion, and thus justifies its derivation monophyletically from the common stem-form Actissa, the different forms of skeleton, on the other hand, cannot all be referred to the same fundamental form. More especially the spherical lattice-shell, from which all the numerous skeletal forms of the Sphaerellaria may be derived, cannot have arisen from the incomplete Beloid skeleton which characterises the Beloidea among the Collodaria. It is probable rather that the formation of the skeleton has taken place independently in those two groups of SPUMELLARIA. From the skeletonless Colloidea, as the common stem-group of the SPUMELLARIA, two different main groups have diverged, on the one hand the Beloidea, whose skeleton consists of separate spicules scattered in the extracapsulum, and on the other hand, the Sphaerellaria, which have formed a simple lattice-sphere around the central capsule ; from this the manifold forms of the remaining SPUMELLARIA may be derived.
163. Descent oj the Sphaerellaria.-The extensive order Sphaerellaria, which includes all SPUMELLARIA with a complete lattice-shell, develops an extraordinary variety of skeletal structures ; these may, nevertheless, all be derived without violence from a common stem-form, or simple spherical lattice-shell, Cenosphaera. The main stem of the order, the extensive suborder Sphaeroidea (Pls. 5-30), is derived immediately from Cenosphaera (p. 61, Pl. 12); three diverging branches of it being represented by the other three suborders, the Prunoidea (Pls. 16, 17, 39, 40) being developed by elongation, and the Discoidea (Pls. 31-48) by shortening of the vertical main axis, whilst the Larcoidea (Pls. 9, 10, 49, 50) have originated by the modification of the spherical lattice-shell into a lentelliptical or triaxial ellipsoidal one. Although the monophyletic derivation of all Sphaerellaria from Cenosphaera is exceedingly probable, the possibility of a polyphyletic origin for the group is by no means excluded. For even in the skeletonless primitive genus of all the SPUMELLARIA, Actissa (as well as in the social Collozoum), there are found, in addition to the usual spherical types, other species (or subgenera, p. 12) whose central capsule is not spherical but a modification of the sphere ; in Actiprunum ellipsoidal ; in Actidiscus lenticular ; in Actilarcus lentelliptical ; if such modified forms of Actissa were to develop their lattice-shells independently, then their form would correspond to that of the central capsule ; and such simple ellipsoidal, discoidal, and lentelliptical lattice-shells might have been the primitive forms of the Prunoidea, Discoidea and Larcoidea.
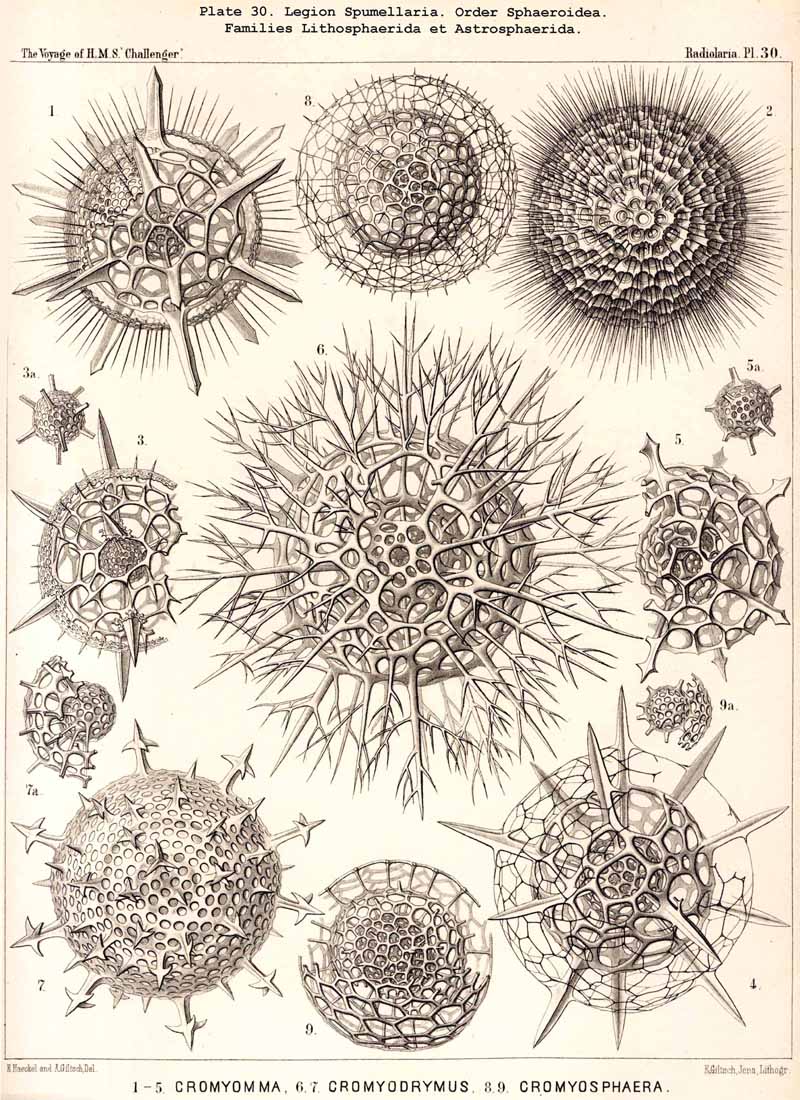
164. Genealogical Tree of the Sphaeroidea.- Cenosphaera, the simplest form of the spherical lattice-shell, may be unhesitatingly regarded as the common stem-form of all the Sphaeroidea (pp. 50-284, Pls. 5-30). Cenosphaera (p. 61, Pl. 12) arose directly from Actissa simply by the silicification of the spherical exoplasmatic network of the sarcodictyum around the central capsule, on the surface of the concentric calymma. From this simple siliceous extracapsular lattice-sphere all other forms of Sphaeroidea have arisen, in the main by the manifold combination of two simple processes, first by the formation of radial spines on the surface of the lattice-sphere, and second, the addition of concentric spherical lattice-shells. Both processes may be utilised as the foundation for a systematic treatment of the Sphaeroidea (compare pp. 52-58).
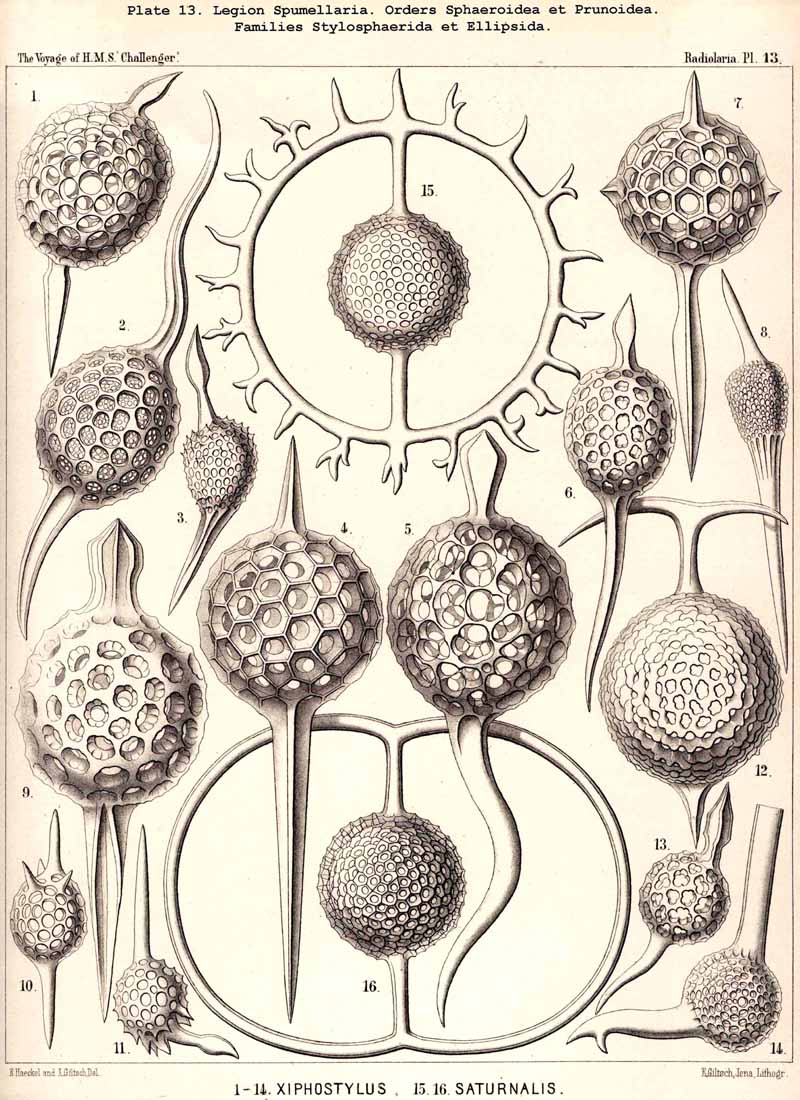
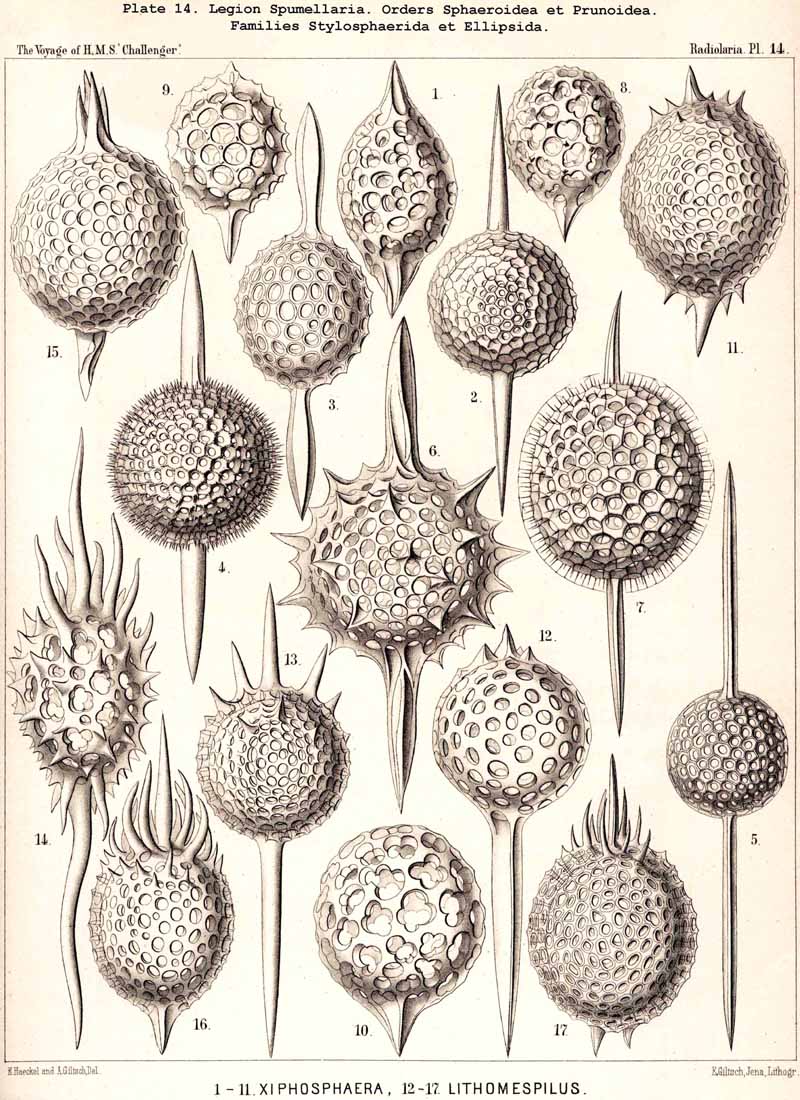
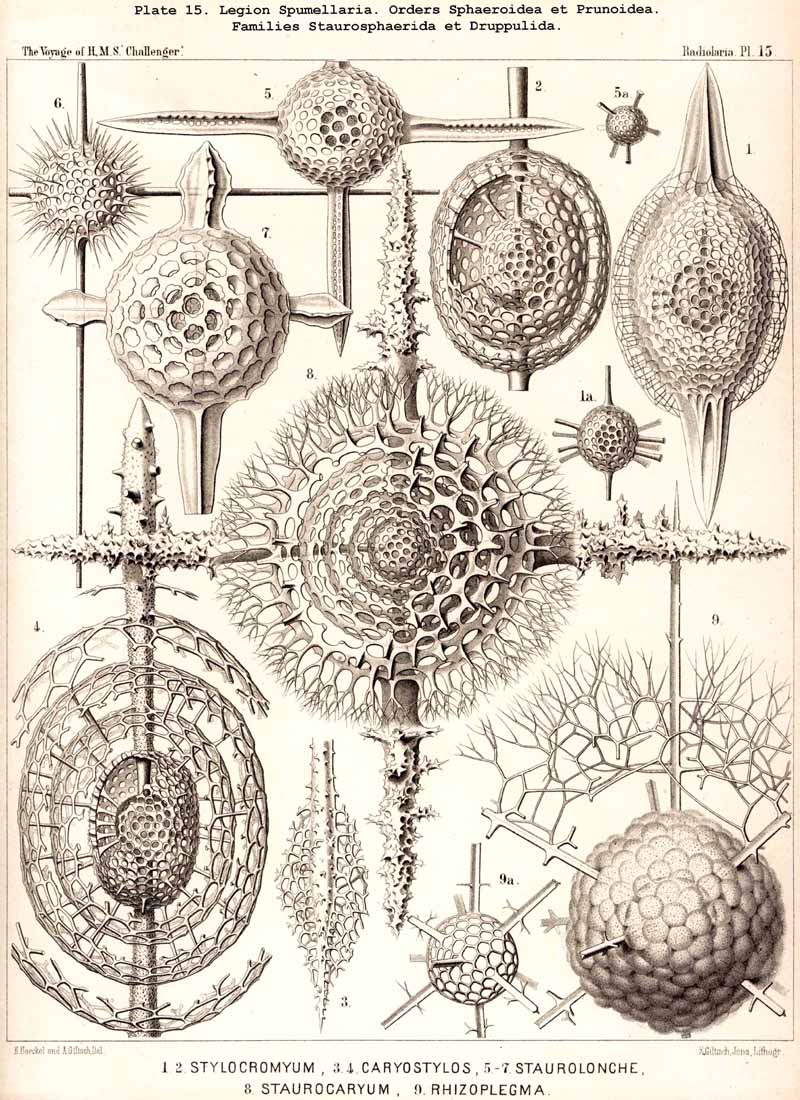
If in the Sphaeroidea the characteristic number and disposition of the radial spines be regarded as the most important heritable peculiarity of the different families, then we have the following natural arrangement :-(1) Liosphaerida, without radial spines ; (2) Cubosphaerida, with six radial spines (opposite in pairs in three axes perpendicular to each other) ; (3) Staurosphaerida, with four radial spines (in two axes crossed at right angles) ; (4) Stylosphaerida, with two opposite radial spines (in the vertical main axis) ; and (5) Astrosphaerida, with numerous regularly or irregularly distributed radial spines (eight to twenty or more). If, on the contrary, more stress be laid upon the number of the concentric lattice-shells, then we have the following artificial grouping :-(1) Monosphaerida, with one simple lattice-sphere: (2) Dyosphaerida, with two concentric lattice-spheres ; (3) Triosphaerida, with three ; (4) Tetrasphaerida, with four ; (5) Polysphaerida, with numerous (five to twenty or more) concentric lattice-shells ; (6) Spongosphaerida, with a spongy spherical shell. In general the former arrangement appears more natural than the latter, since the number of primary radial spines, which grow out from the primary lattice-sphere, determines their ground-form from the outset, whatever may be the number of secondarily added shells. Strictly speaking, according to the view adopted, those Liosphaerida which have several shells, on the outer surface of which there are no radial spines, ought to be classified according to the number and arrangement of their internal radial connecting beams and distributed among the other families. The practical application of this correct principle meets, however, with great difficulties. Also in many cases the phylogenetic relations of the different Sphaeroidea are more complicated than would appear from both these classificatory principles. In general their phylogeny will quite correspond with their ontogeny, since from the innermost first formed lattice-shell (primary medullary shell) a number of radial spines arises, and upon these the secondary shells are formed from within outwards.
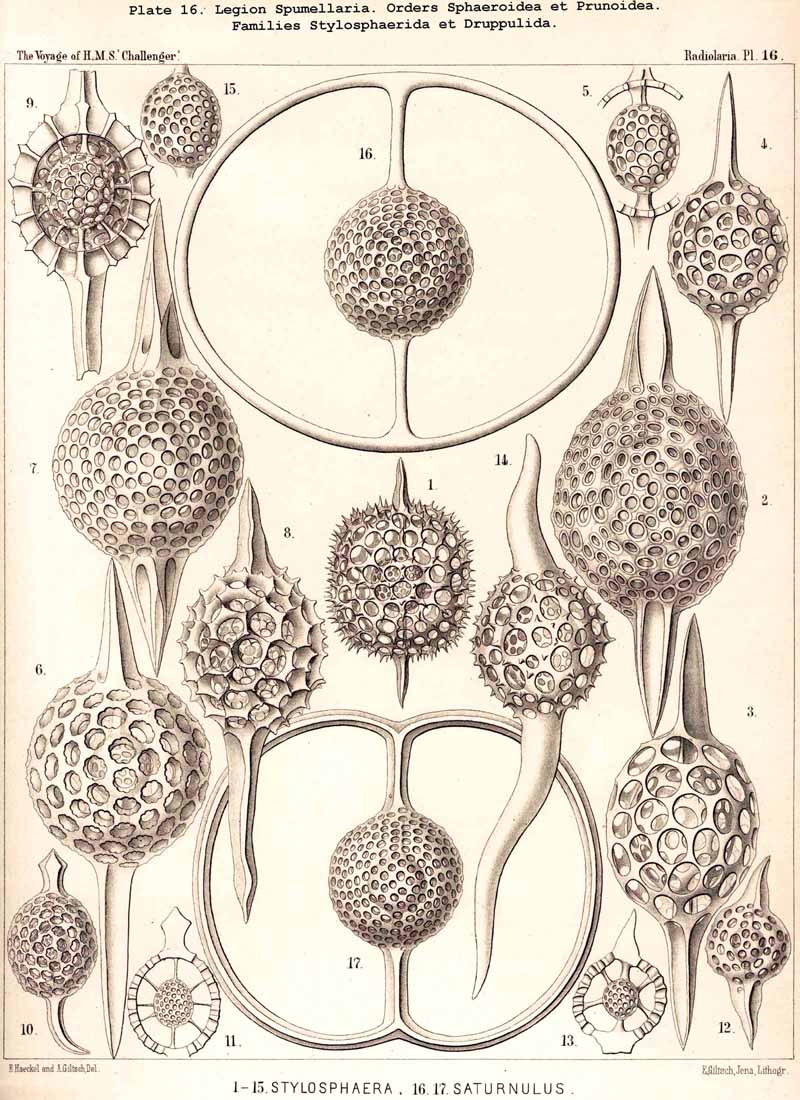
165. Genealogical Tree of the Prunoidea.-The suborder Prunoidea is very closely related to the Sphaeroidea, and is distinguished from it by the elongation of one axis ; from the simple lattice-sphere (Cenosphaera) is developed a latticed ellipsoid (Cenellipsis, Pl. 39, fig. 1). The development of this vertical isopolar main axis is foreshadowed even among the Sphaeroidea, in that family in which two opposite radial spines grow out of the primary lattice-sphere at the two poles of the vertical main axis (Stylosphaerida, Pls. 13, 14). These latter pass over without any sharp boundary into those forms of Prunoidea whose ellipsoidal lattice-shell bears two opposite main spines (Stylatractida, Pls. 15, 16). Other very intimate relationships between the Sphaeroidea and Prunoidea are indicated in certain of the latter by the fact that of the two concentric lattice-shells the inner (medullary) shell is spherical, the outer (cortical) shell ellipsoidal (Pl.39, figs. 3, 7, 8, 14, 19); often three concentric lattice-shells are present, of which the two inner are spherical intracapsular medullary shells, whilst the outer is an extracapsular cortical shell, ellipsoidal or cylindrical in form (Pl. 39, figs. 4, 12, 17, 18). Owing to the manifold nature of these phylogenetical relations and the variety of their combinations, the derivation of the individual Prunoidea from the Sphaeroidea is rendered very difficult ; in addition to which it is possible that the simplest Prunoidea (C1enellipsis, Ellipsidium) have been directly developed from the skeletonless Actiprunum (a form of Actissa with ellipsoidal central capsule, p. 14) by the excretion of a simple ellipsoidal lattice-shell on the surface of their calymma.
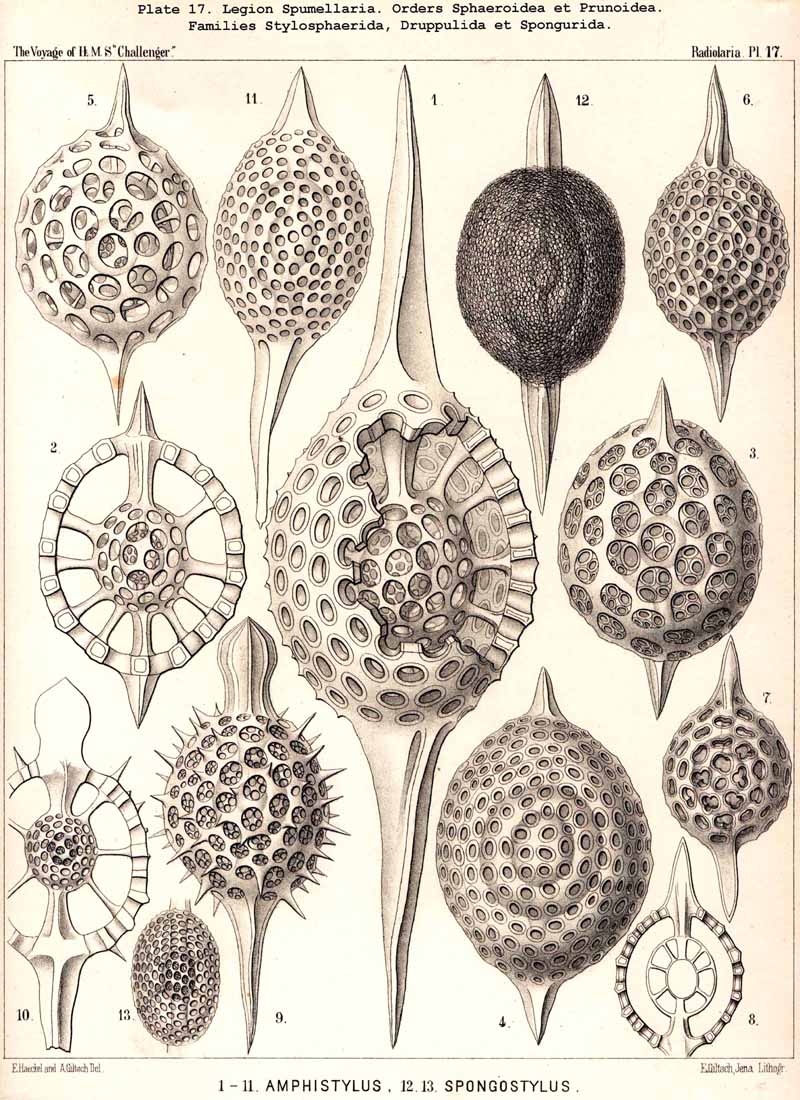
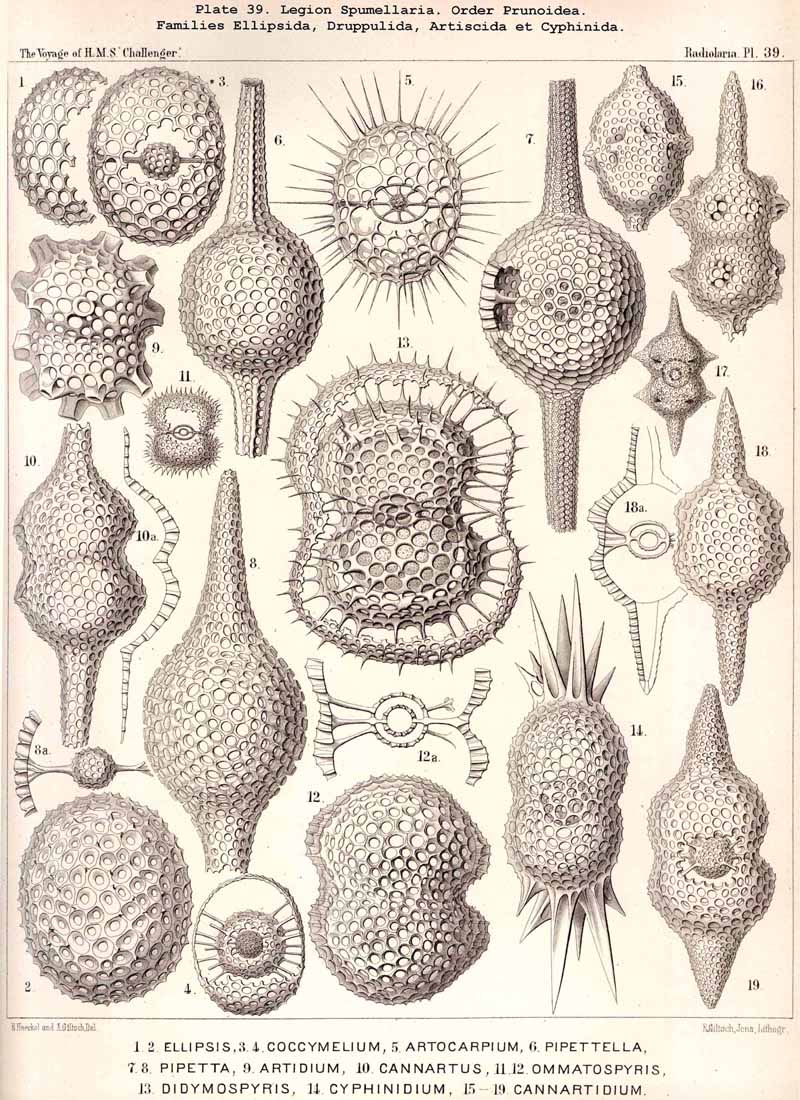
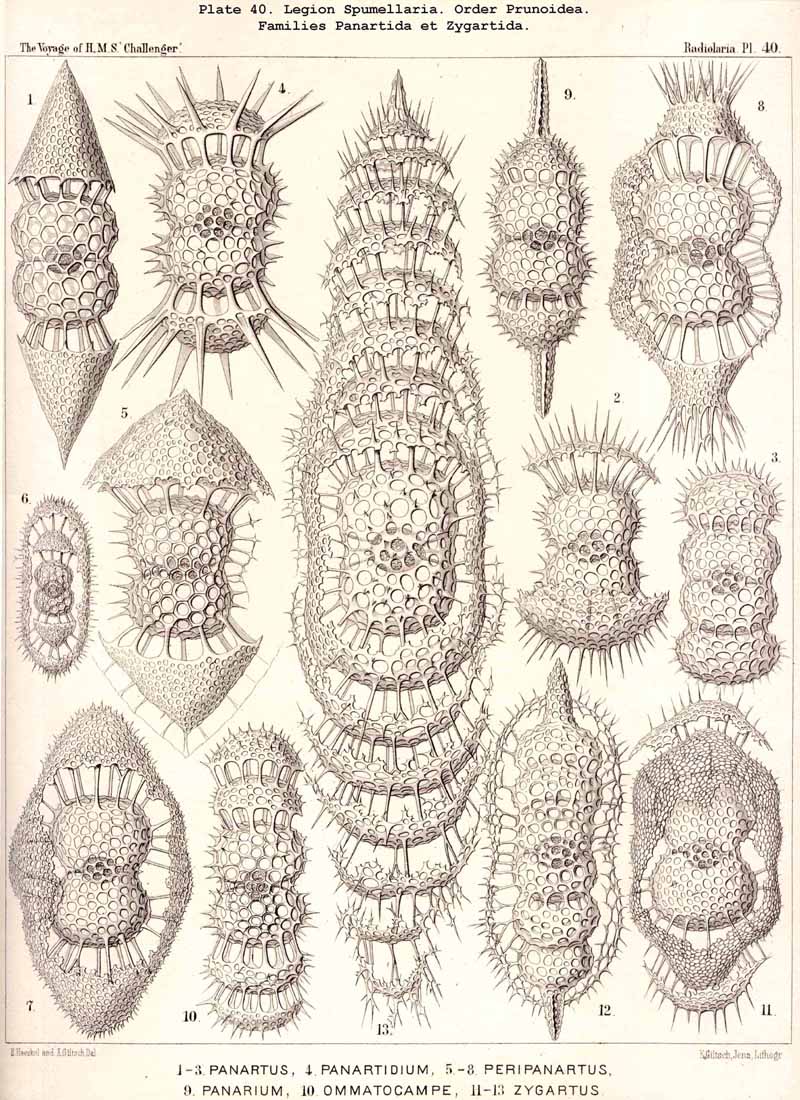
The phylogeny of the Prunoidea is especially complicated by the formation of peculiar transverse constrictions, perpendicular to the longitudinal axis. They are wanting only in the Monoprunida (Ellipsida, Druppulida, and Spongurida) ; the Dyoprunida (Artiscida and Cyphinida, Pl. 39, figs. 9-19) possess only one such constriction (in the equatorial plane) ; the Polyprunida, on the other hand, have three, five, or more parallel constrictions (Panartida and Zygartida, Pl. 40). The chambers, which are separated off by these constrictions, may be regarded as polar sections of incomplete cortical shells.
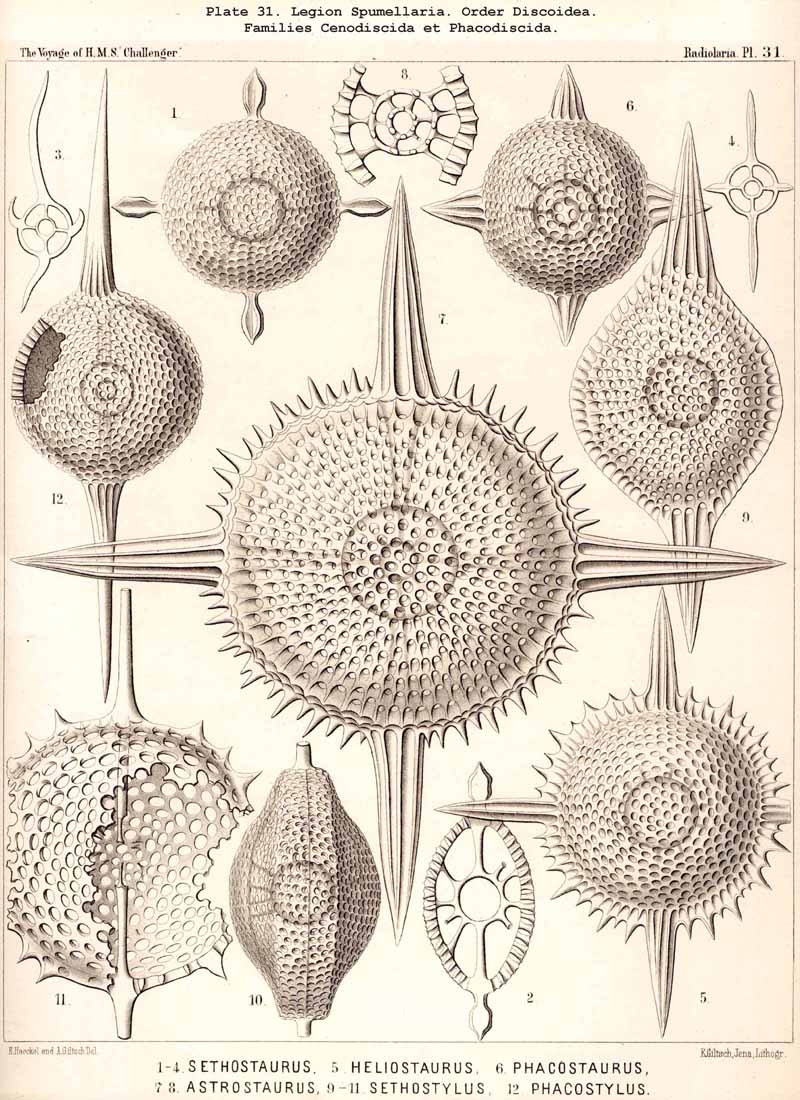
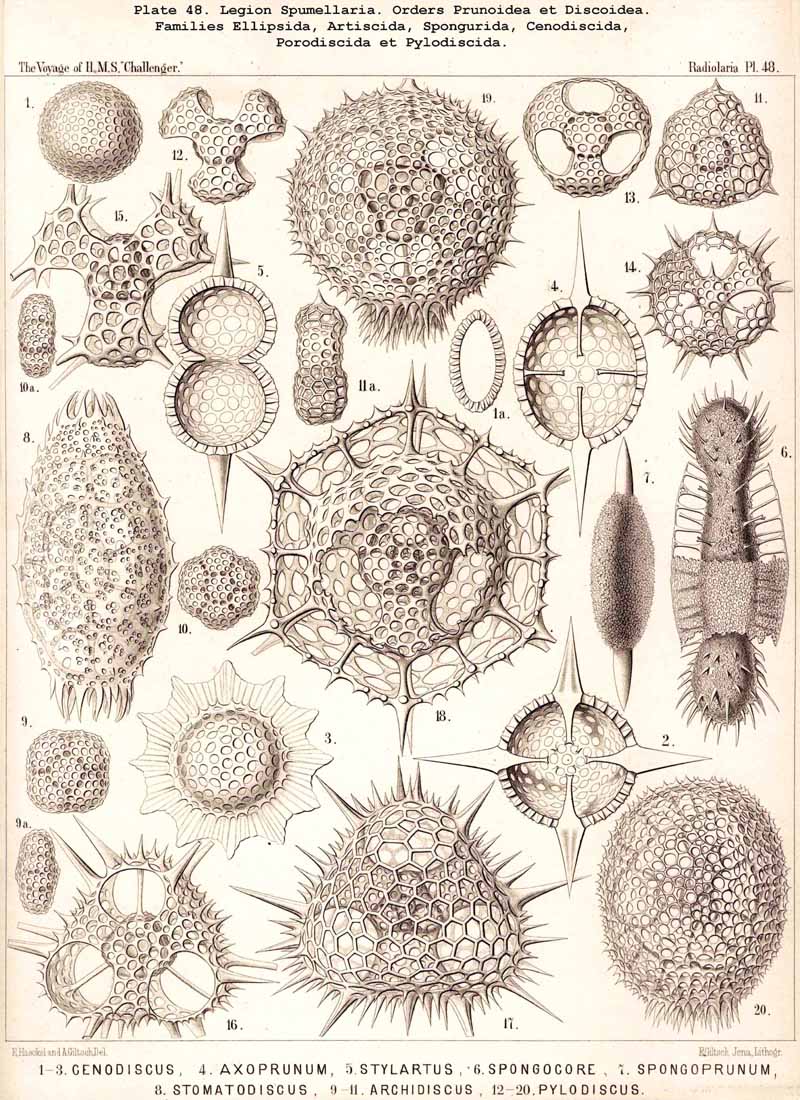
166. Genealogical Tree of the Discoidea.-The suborder Discoidea is closely related to the Sphaeroidea, but separated from it by shortening of one axis ; from a simple lattice-sphere (Cenosphaera) a latticed lens or flattened spheroid is developed, whose circular equatorial plane is larger than any other section (Cenodiscus, Pl. 48, fig. 1). The formation of this horizontal equatorial plane is perhaps indicated in that family of Sphaeroidea in which four crossed radial spines, lying in one plane, are developed (Staurosphaerida, Pls. 15, 31, 42). The morphological and phylogenetical relations of the Discoidea to the Sphaeroidea are precisely the converse of those of the Prunoidea ; in the latter the vertical axis appears longer, in the former shorter than any other axis of the body. The Discoidea are probably polyphyletic, having originated from several different groups of Sphaeroidea ; at least two essentially different main groups may be distinguished among them ; of these the one is characterised by the formation of a large extracapsular lenticular cortical shell (Phacodiscaria), whilst in the other this typical "Phacoid shell" or lattice-lens is wanting (Cycbodiscaria, compare pp. 403-409).
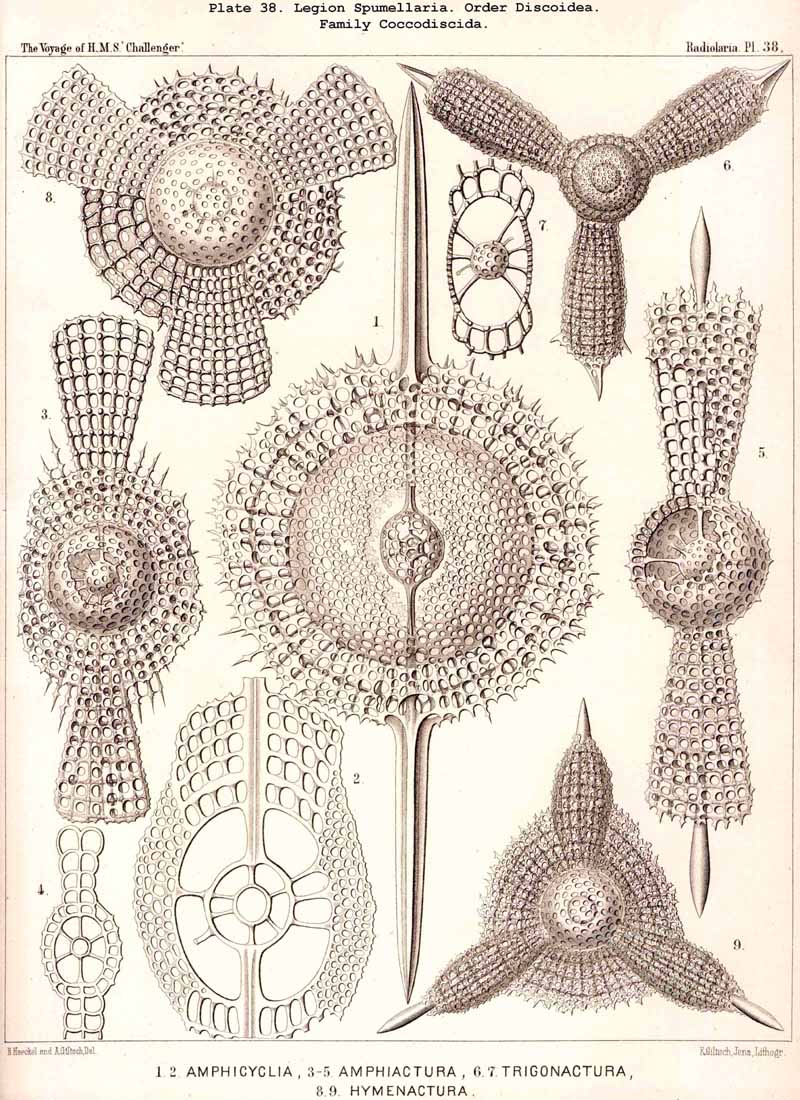
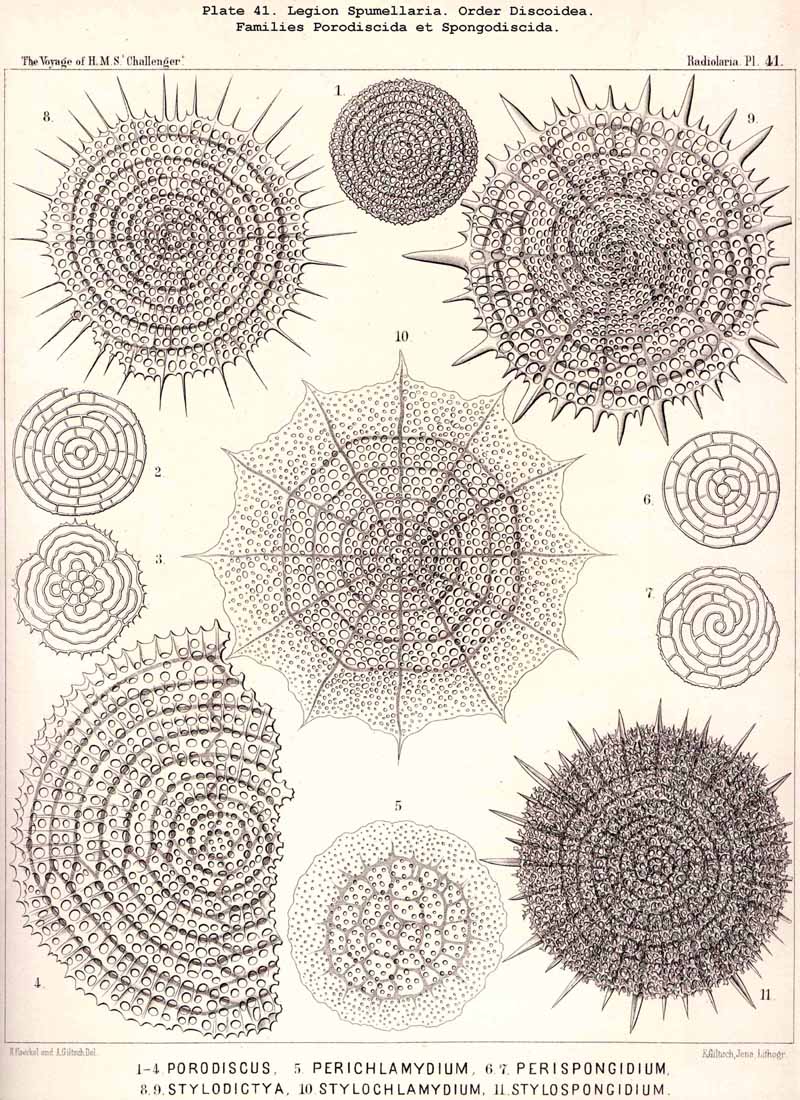
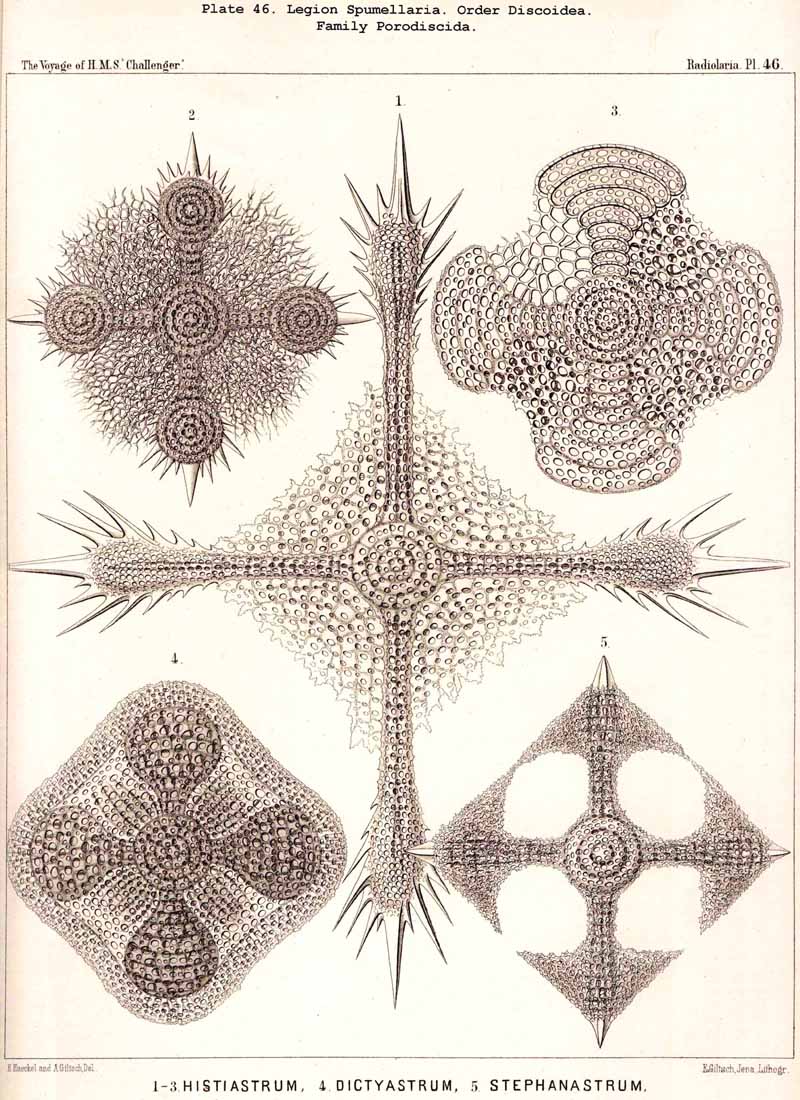
The Phacodiscida (Pls. 31-35) perhaps constitute the primitive group of the Phacodiscaria, their lenticular or Phacoid cortical shell being connected by radial bars with one or two concentric spherical medullary shells ; they may have originated directly from the Dyosphaerida or Triosphaerida by flattening of the spheroidal cortical shell. From the Phacodiscida the Cenodiscida (if indeed they be not the primitive stem-form) have been developed by retrogression and loss of those medullary shells. The Coccodiscida (Pls. 36,37, 38), on the other hand, have been developed from the Phacodiscida by the addition of concentric rings of chambers, which may be regarded as incom plete cortical shells, only the equatorial portion of which is developed. Perhaps the Porodiscida, the primitive group of the Cyclodiscaria, have arisen in a similar way ; they lack, however, the typical Phacoid shell, the concentric rings of chambers being directly applied to a small spherical medullary shell in the equatorial plane (Pls. 41-46). If those rings from the commencement be interruped by three interradial gaps (gates) the family Pylodiscida arises (Pl. 38, figs. 6-20). If, on the contrary, the concentric radially divided chambers of the Porodiscida become quite irregular and spongy, they pass over into the Spongodiscida (Pls. 46, 47). It is not, however, impossible that part of the Discoidea (especially the Cenodiscida) have originated directly from skeleton-less Collodaria with a lenticular central capsule, such as are found in a subgenus of Actissa (Actidiscus, p. 15).
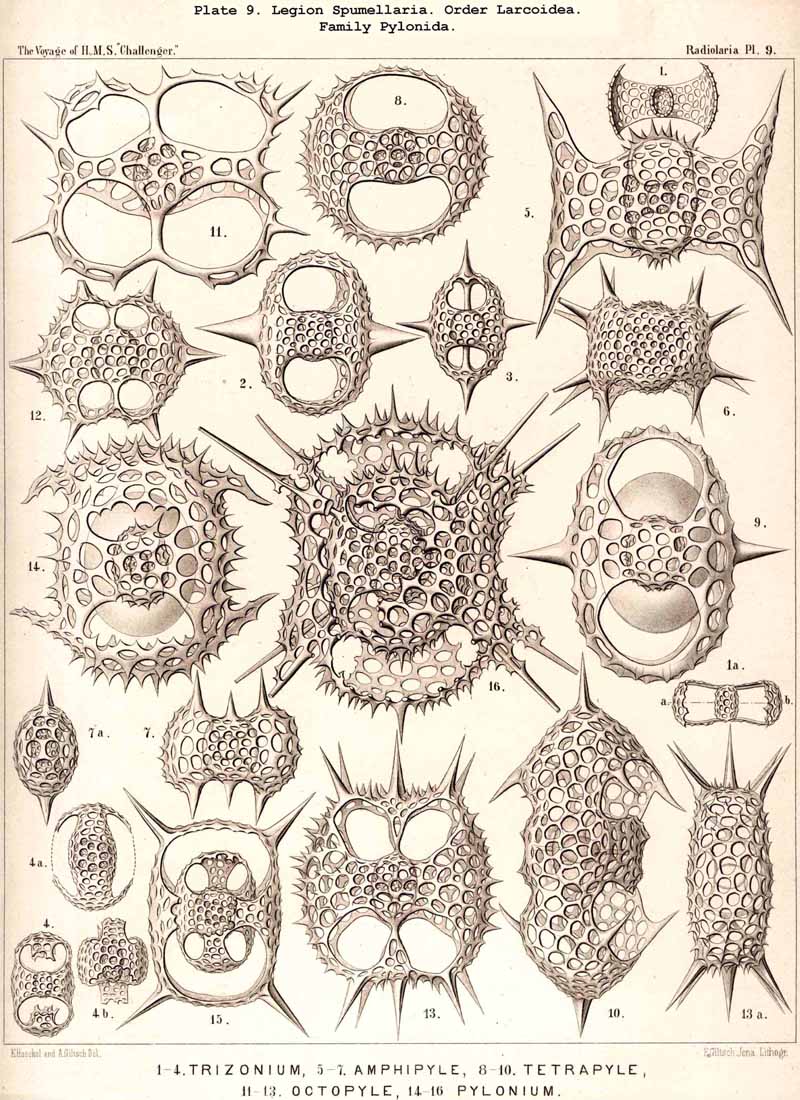
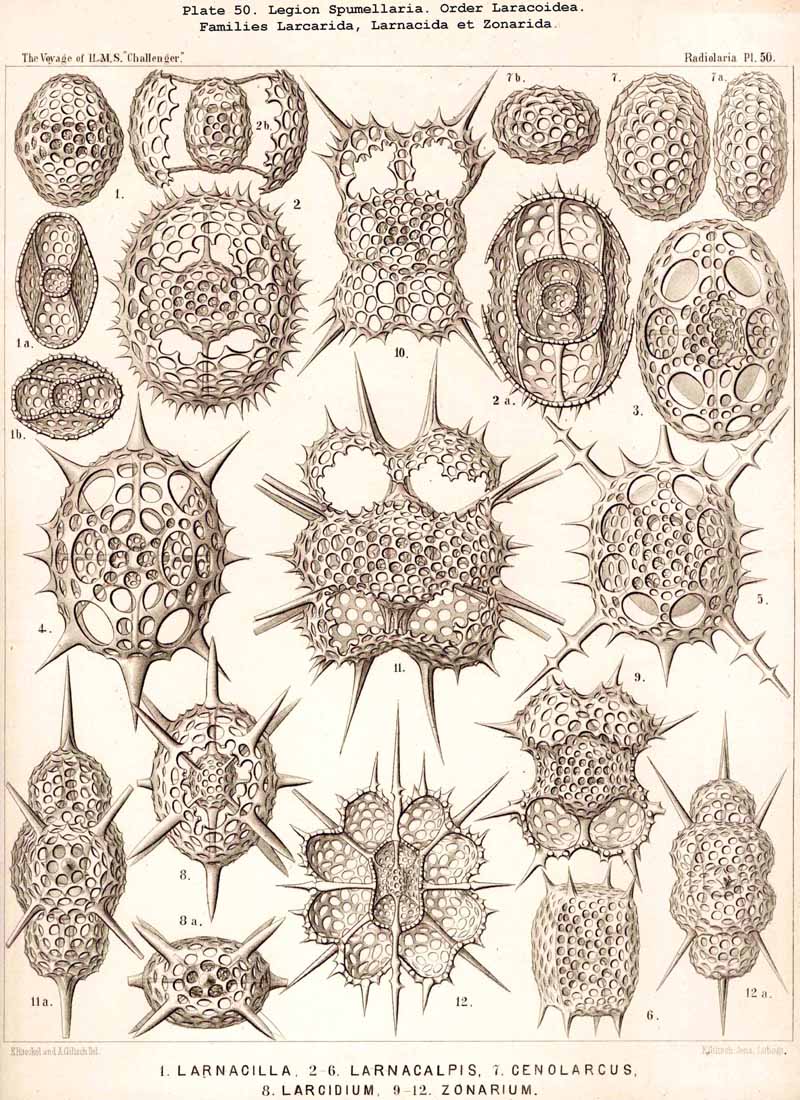
167. Genealogical Tree of the Larcoidea.-The suborder Larcoidea presents in the structure, composition, and development of its variously formed lattice-shells much more complicated relations than the other Sphaerellaria; it is essentially distinguished from them by the characteristic ground-form of its lattice-shells, which is a "lentellipsis" or a triaxial ellipsoid (also the ground-form of the rhombic crystallographic system, the rhombic octahedron). Hence all parts of the body are regularly disposed with respect to three different dimensive axes ; all three axes, perpendicular one to another, are isopolar but of different lengths ; the longest is the vertical main axis, the mean the horizontal frontal axis, the shortest the horizontal sagittal axis. In the great majority of the Larcoidea the lentelliptical ground-form is indicated in the central capsule, even when it is not at once obvious in the skeleton. Since such lentelliptical central capsules are developed even in Actissa (Actilarcus, p. 16), it is possible that the simplest Larcoidea may have arisen directly from these by deposition of a simple lentelliptical lattice-shell in the sarcodictyum, on the surface of the calymma (Cenolarcus, Pl. 50, fig. 7). It is more probable, however, that these simplest forms (Cenolarcus, Larcarium) have been developed from the simplest Sphaeroidea (Cenosphaera), by the spherical body growing unequally in the three dimensions of space. It appears especially likely from a study of the concentrically disposed lattice-shells of some Larcoidea (Coccolarcus, Larcidium, Pl. 50, fig. 8), in which the inner medullary shell is spherical, the outer cortical shell more or less elliptical. In the great majority of Larcoidea the batter arises in quite a peculiar manner, three broad lattice-zones, which are developed in three planes at right angles to each other, growing out from a small spherical or lentelliptical medullary shell, Trtzonium, Larnacilla (compare pp. 600, 615, 628, &c.).
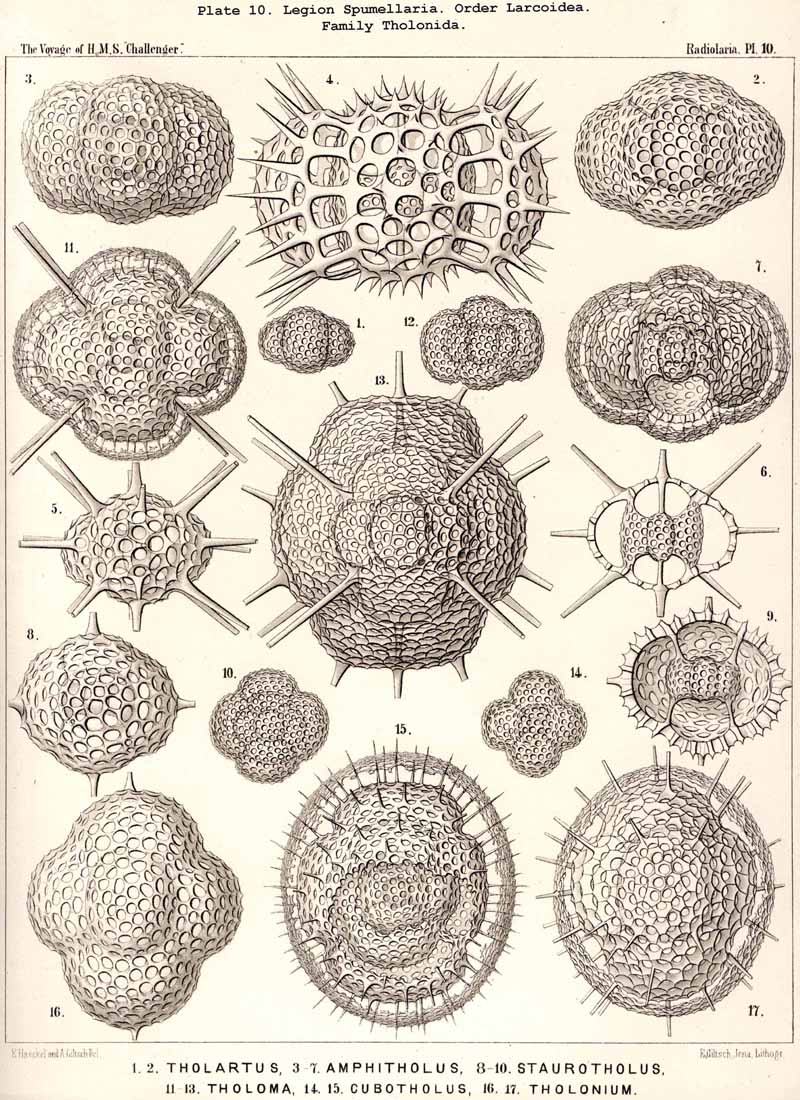
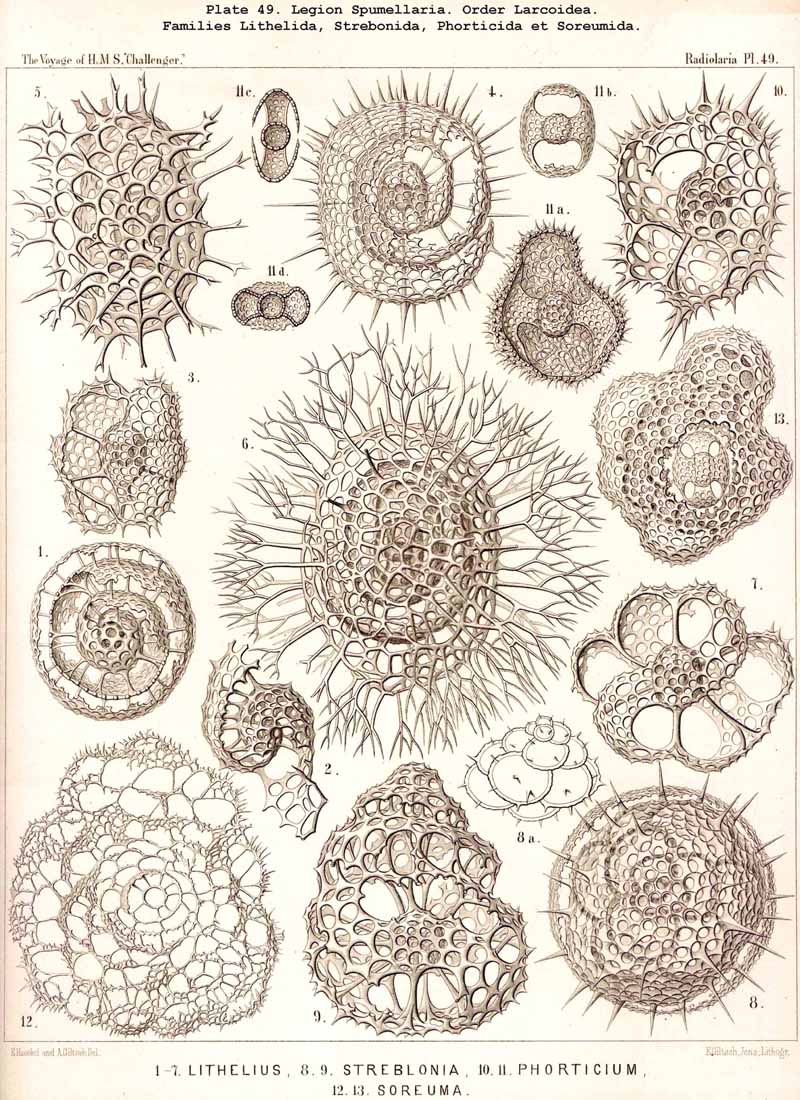
The trizonal Larnacilla-shell commences by the formation of a transverse girdle, by the union of two lateral latticed processes, which spring right and left in the equatorial plane from the poles of the frontal axis of a lentelhiptical medullary shell (Mionozonium, p. 633, Pl. 9, fig. 1). This is followed by a second lateral girdle, which lies in the frontal plane and proceeds from its lateral poles (Dizonium, p. 634, Pl. 9, figs. 2, 3). Finally the sagittal girdle is formed, lying in the sagittal plane and arising from the lateral girdle on the two poles of the main axis (Trizonium, p. 637, Pl. 9, fig. 4). Whilst the gaps between the three zones of this trizonal shell remain open in the Pybonida, in Larnacilla, the important primitive form of the Larnacida, they are closed by lattice-work (Pl. 50, figs. 3-8). From this trizonal Larnacilla-shell the great majority of Larcoid shells may be derived. Such a system of zones may be repeated (Diplozonaria) or even developed a third time (Triplozonaria, p. 632). In most Larcoidea the zones are secondarily connected by lattice-work. In the Tholonida (Pl. 10) each of the two opposite latticed wings of a zone becomes a closed dome. In the Zonarida (Pl. 50 figs. 9-12) these domes are partially or wholly bisected by constrictions or latticed septa which are developed in the three dimensive planes. The Lithelida (Pl. 49, figs. 1-7) are characterised by the fact that one of each pair of opposite latticed processes (or half zones) grows more strongly than the other, and that the larger completely embraces the smaller so as to form a complicated spiral. Whilst in this case the spiral lies in a plane, in the Streblonida (Pl. 49, figs. 8, 9) it becomes turbinoid like a gastropod shell and forms an ascending spiral. Finally, two small families of Larcoidea are characterised by quite irregular growth (a very rare occurrence among the Radiolaria) ; these are the simple-chambered Phorticida (Pl. 49, figs. 10, 11), and the many chambered Soreumida (Pl. 49, figs. 12, 13). The phylogenetic relationship of these families of Larcoidea is probably very complicated and demands closer investigation (compare pp. 599-604).
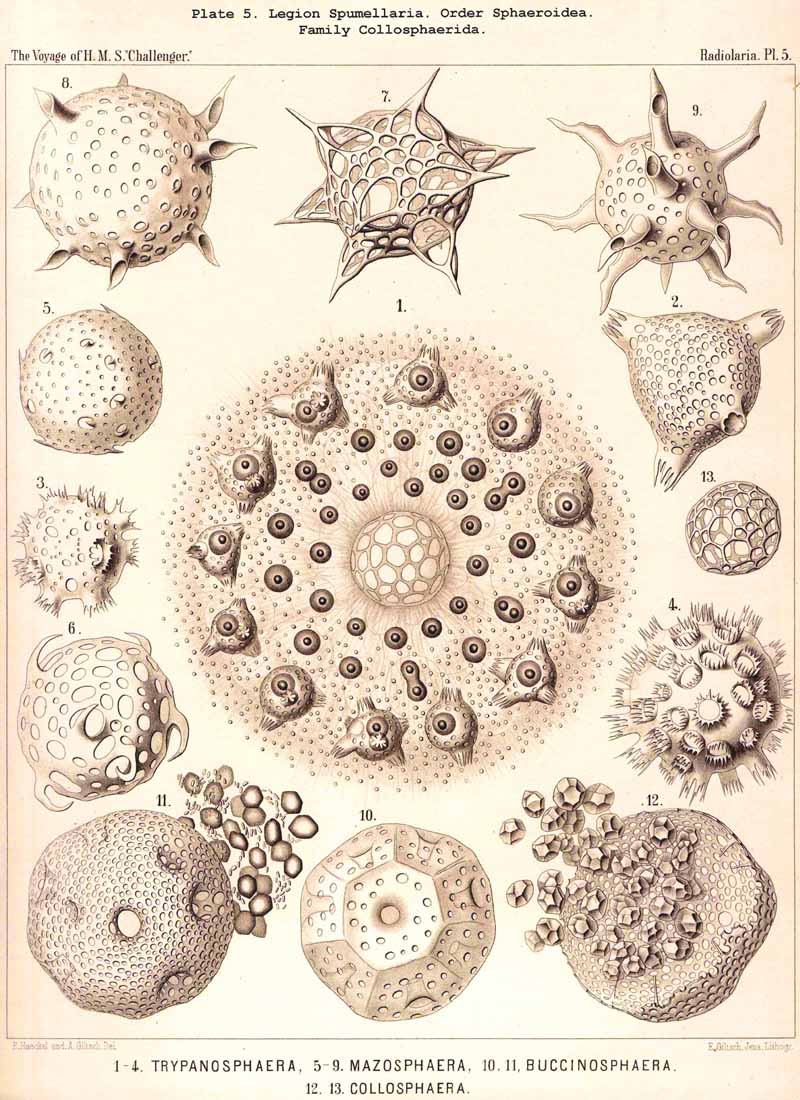
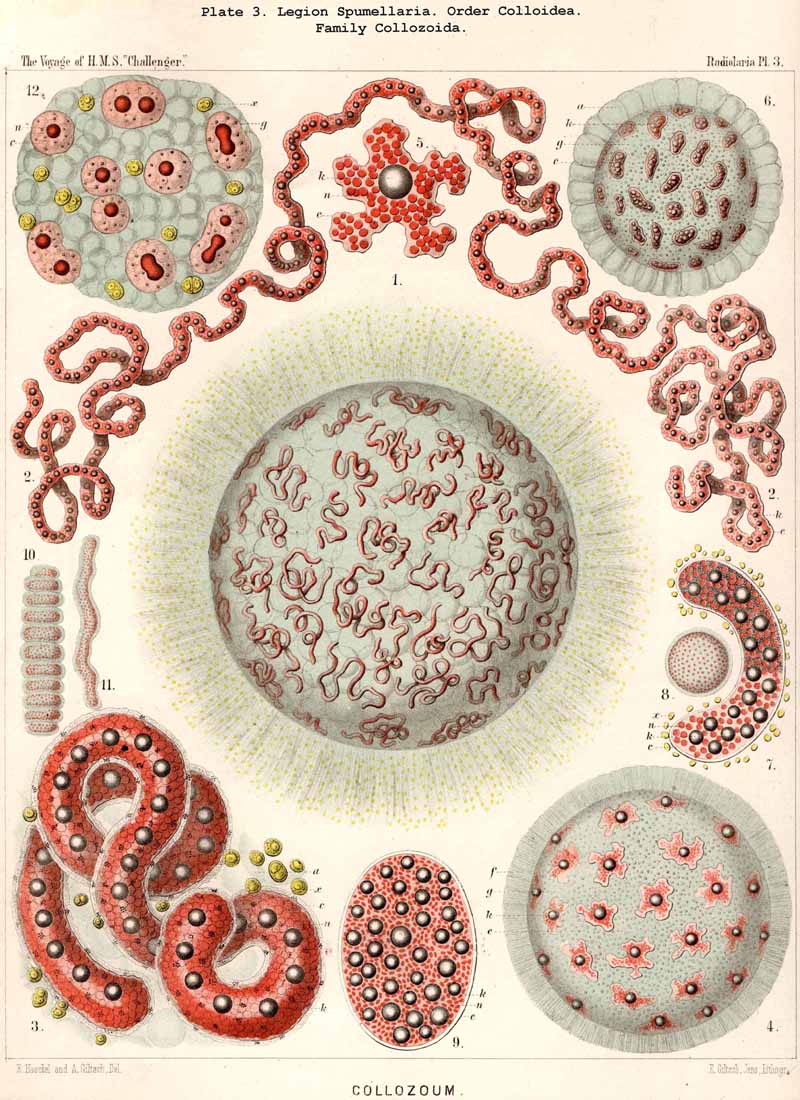
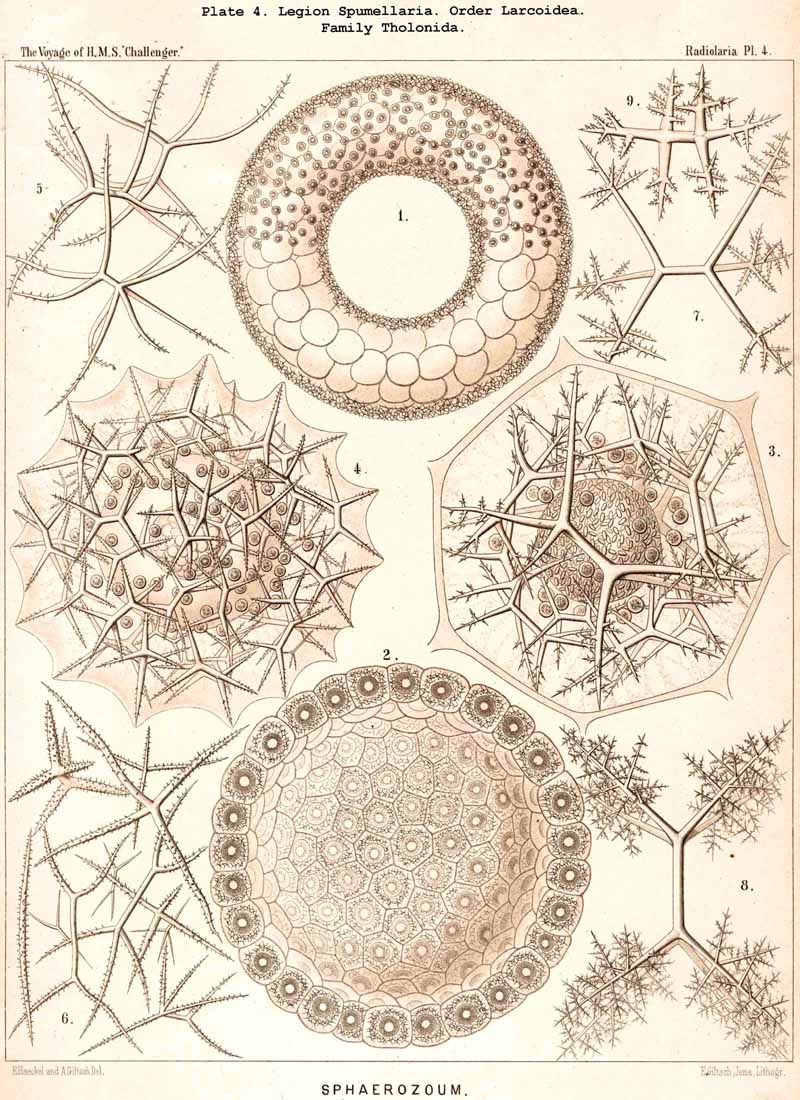
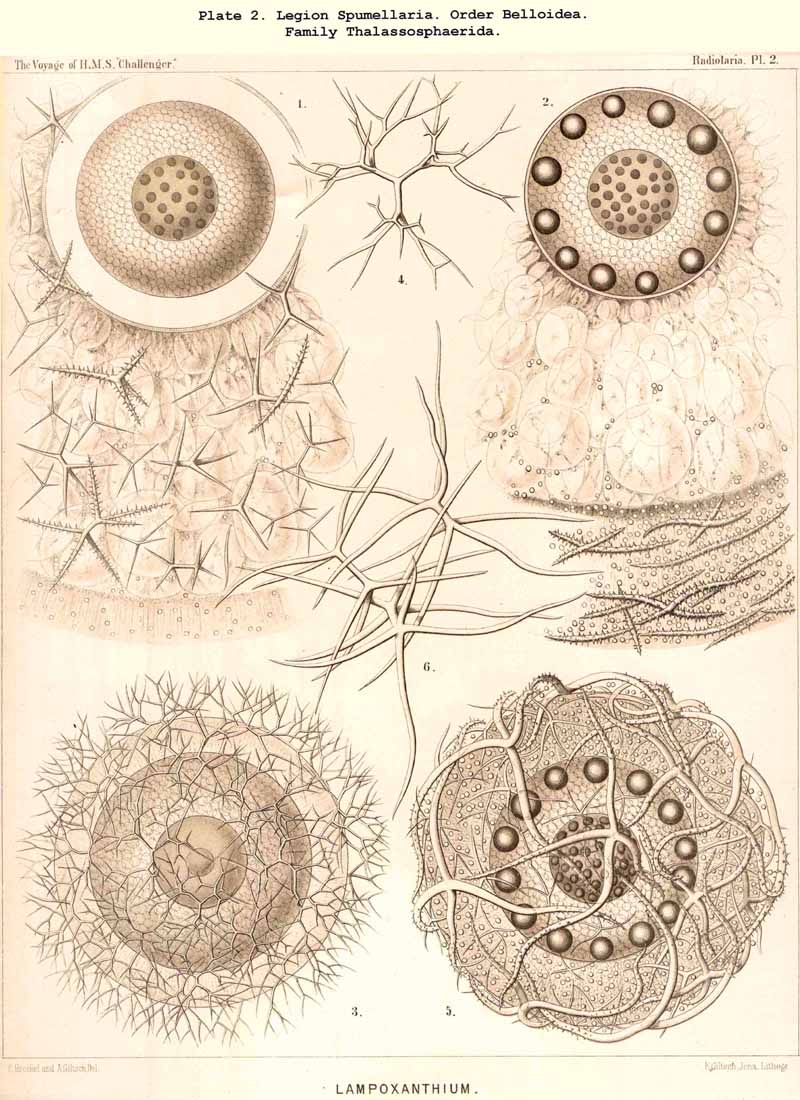
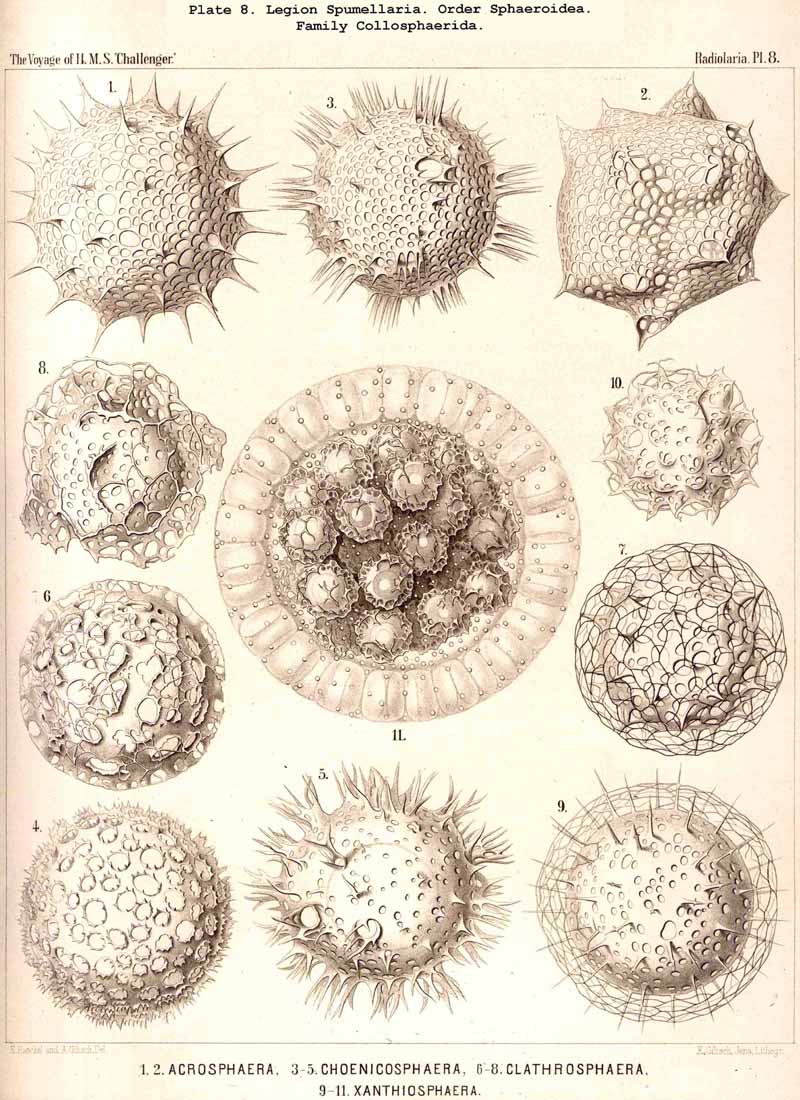
168. Descent of the Polycyttaria.-The polyzootic or colonial Radiolaria, which we unite in the group Polycyttaria (sometimes known as "Sphaerozoea"), belong without doubt to the legion SPUMELLARIA, for they possess all the peculiarities by which these PERIPYLEA are distinguished from the other legions of the Radiolaria. Only the morphological position of the Polycyttaria in that legion, and their phylogenetic relation to the monozootic or solitary SPUMELLARIA, can be variously interpreted. The three families which we distinguish among the Polycyttaria are so closely related to three different families of the Monocyttaria, that they may be directly derived from them by the formation of colonies. According to this triphyletic hypothesis the social skeletonless Collozoida (Pl. 3) would be descended from the solitary Thalassicollida (Pl. 1), the polyzootic Sphaerozoida with a Beboid skeleton (Pl. 4) from the monozootic Thalassosphaerida (Pl. 2), and the colonial Collosphaerida with a Sphaeroid skeleton (Pls. 5-8) from the solitary Ethmosphaerida (Pl. 12, &c.). Many species of monozootic and polyzootic forms in all three groups are so alike that they can only be distinguished by the fact that the one series are colonial, the others solitary. On the other hand, there are some reasons which would justify a monophyletic hypothesis for the Polycyttaria, e.g., the precocious nuclear division ; in this case it would be most natural to hold that the Sphaerozoida and Collosphaerida have arisen as two diverging branches from the Collozoida, whilst the batter are nothing else than colonial Thalassicollida.
169. Phylogeny of the Acantharia.-The legion ACANTHARIA or ACTIPYLEA is distinguished by its peculiar acanthin skeleton, which develops centrogenously, as well as by the disposition in groups of the pores in its central capsule, and its excentric usually precocious nucleus ; it is thus so different from all other Radiolaria as undoubtedly to furnish, phylogenetically considered, an independent stem (§ 7). This stem is only connected at the root by Actinelius with the primitive form of the SPUMELLARIA, Actissa. The stem is monophyletic, since all the forms belonging to it may be derived without violence from Actinelius as a common primitive form.
170. Origin of the Acantharia.-The genus Actinelius (p. 730, Pl. 129, fig. 1), which may naturally be regarded as the common primitive form of all ACANTHARIA, possesses a spherical central capsule, which in consequence of the early division of the nucleus (§ 63), encloses numerous small nuclei ; from its centre arise many simple radial spines of equal size, which penetrate the central capsule. A large number of radial pseudopodia issue between the spines from the sarcomatrix which surrounds the capsule. Actinelius may have been directly derived from Actissa, the common stem-form of all Radiolaria, by the division of the pseudopodia into two groups, myxopodia, which remained soft, and axopodia, which became firm (§ 95A). As the latter became changed into strong acanthin rods, and touched each other in the centre, they forced the nucleus from its originally central position and brought about its early division. Actinelius is also of all Radiolaria the form which, next to Actissa, most nearly approaches the Heliozoa. If the stiff axial threads of Actinosphaerium be conceived of as partially converted into acanthin spines, and its nucleated medullary substance as separated from the alveolar cortical layer by a membrane (central capsule), then Actinelius would be produced.
171. Hypothetical Genealogical Tree of the Acantharia.
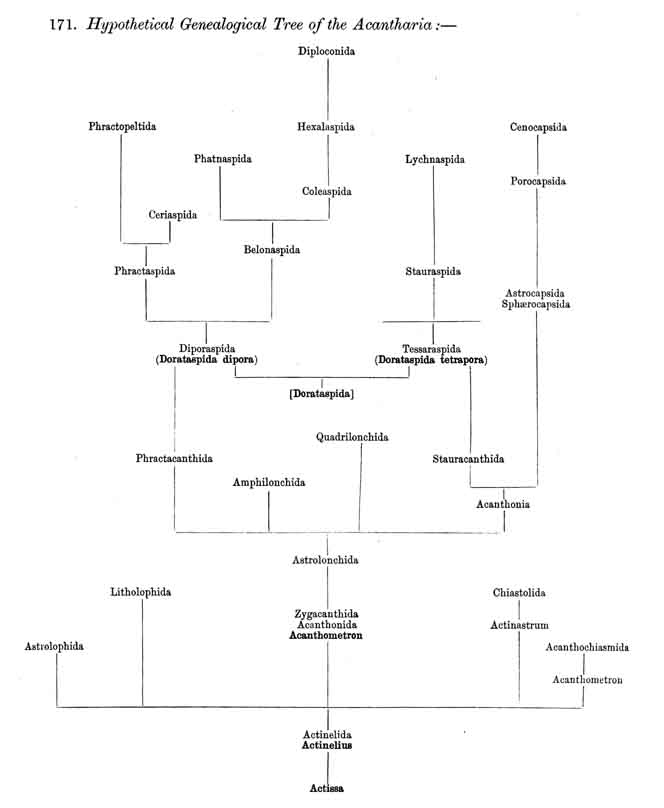 |
172. Adelacantha and Icosacantha.-The numerous forms of ACANTHARIA, here disposed in twelve families and sixy-five genera, may be divided phylogenetically into two main groups of very different extent -Adelacantha and Icosacantha. The more primitive group, Adelacantha, have an indefinite and variable number of radial spines, which are always quite simple in form and usually irregularly distributed ; this main division includes only the one order Actinelida, with six genera, among which is Actinelius, the common stem-form of all the ACANTHARIA. The more recent group, Icosacantha, includes all the other ACANTHARIA (fifty-nine genera), and is very markedly distinguishied from the Adelacantha by the fact that the radial spines are always twenty in number, and arranged according to Müller's law (compare pp. 717-725, and § 110). Since this regular disposition (in five alternating zones each of four spines) has been retained by inheritance in the whole of the Icosacantha, it is probable that this large group has been developed monophyletically from a twig of the Adelacantha ; Actinastrum (p. 732) and Chiastolus (p. 738) still present connecting links between the former and the latter, between Actinelius and Acanthometron.
173. Acanthonida and Acanthophracta. -The extensive main division Icosacantha (§ 110), which embraces all ACANTHARIA with twenty radial spines, disposed according to Müller's law, may be subdivided into two large groups or orders :-the Acanthonida (p. 740, Pls. 130, 131, 132) and the Acanthophracta (p. 791, Pls. 133-140). The latter possess a complete extracapsular lattice-shell, which the former have not. The more recent Acanthophracta may be derived phylogenetically from the more primitive Acanthonida simply by the development of this lattice-shell, with which process are usually (perhaps always) connected certain alterations in the mahacoma, e.g., degeneration of the myophriscs (§ 96). The most primitive form of all Icosacantha is the genus Acanthornetron (p. 324), in which all the twenty acanthin spines are of the simplest constitution and of equal dimensions.
174. Differentiation of the Acanthonida.-The order Acanthonida, which embraces all Icosacantha which have no complete lattice-shell, divides early into three main branches, the three families Astrolonchida, Quadrilonchida, and Amphilonchida (p. 727, Pls. 130, 131, 132). The first of these constitutes the common stem-group from which the other two as well as the whole group Acanthophracta have been developed ; the common steni-form of all is Acanthometron (§ 173). All the Astrolonchida (p. 740, Pl. 130) have twenty radial spines of equal size and similar form. On the other hand, in the Quadrilonchida (p. 766, Pl. 131) the four equatorial spines differ from the others in size and sometimes also in form. In the Amphilonchida (p. 781, Pl. 132) two opposite equatorial spines (hying in the hydrotomical axis) are much larger than the other eighteen and of a different shape. Of the three families of the Acanthonida the most important is the primitive group Astrolonchida, for from this the various stem-forms of the Acanthophracta arise. They are subdivided according to the formation of the spines into three subfamihies: the Zygacanthida, with simple spines without apophyses (or transverse processes) ; the Phractacanthida, with two opposite apophyses on each radial spine, and the Stauracanthida, with four crossed apophyses on each radial spine. The three genera of the Zygacanthida represent the stem-forms of the three families, since the radial spines in Acanthometron (the most primitive form of Acanthonida) are cylindrical, in Zygacantha two-edged, and in Acanthonia four-edged (p. 741).
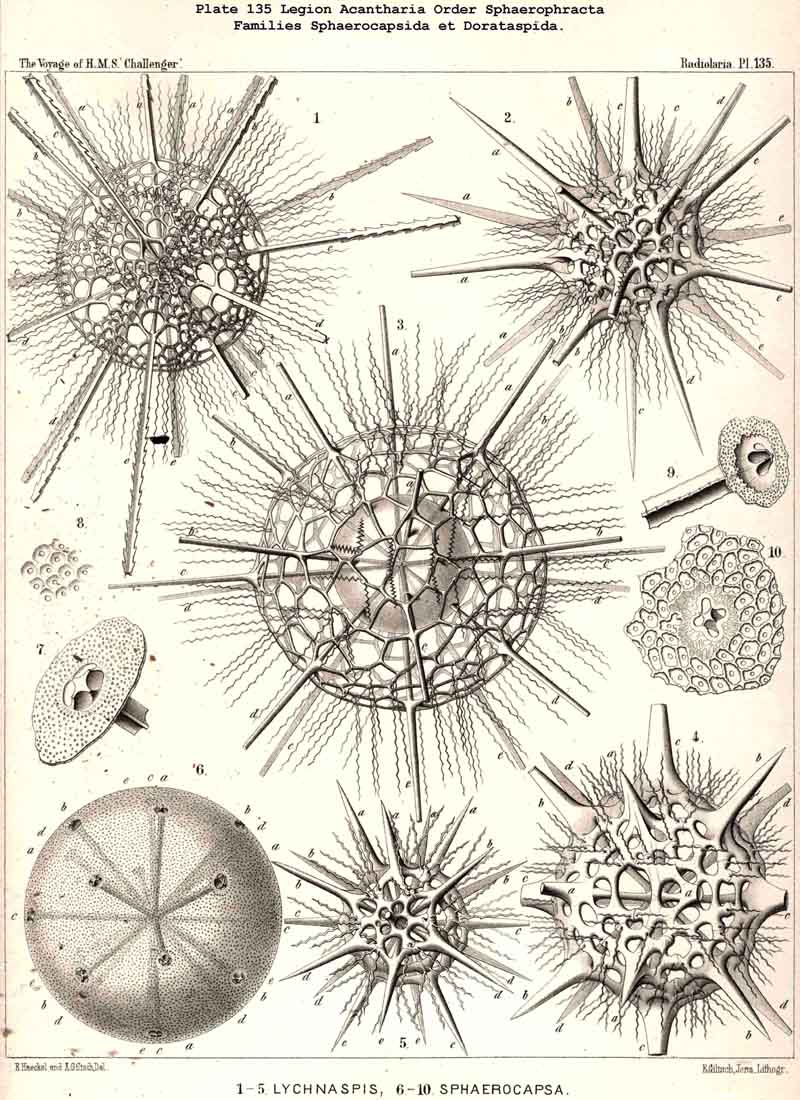
175. Capsophracta and Cladophracta.-The extensive order Acanthophracta, which embraces all ACANTHARIA with a complete lattice-shell, is polyphyletic, its main subdivisions have been developed independently from different branches of the Acanthonida. The whole order may be divided directly into two main groups, the Capsophracta and Cladophracta (p. 793), which differ in the structure and the origin of their lattice-shell. The group (or suborder) Capsophracta includes only the single family Sphaerocapsida (p. 795, Pl. 133, figs. 7-11 ; Pl. 135, figs. 6-10); the lattice-shell arises independently of the twenty radial spines, being made up like a pavement of innumerable small acanthin plates, united by a kind of cement ; each plate being perforated by a fine pore. In addition twenty larger main pores (or groups of four pores each) are present, corresponding to the twenty radial spines ; these are always equal, quadrangular prismatic, without transverse processes as in Acanthonia. In the Cladophracta, which include the five remaining families of the Acanthophracta, the structure and origin of the lattice-shell are quite different ; the lattice-shell is here made up of the branches of the transverse processes, which radiate tangentially from the twenty. radial spines and are only united secondarily.
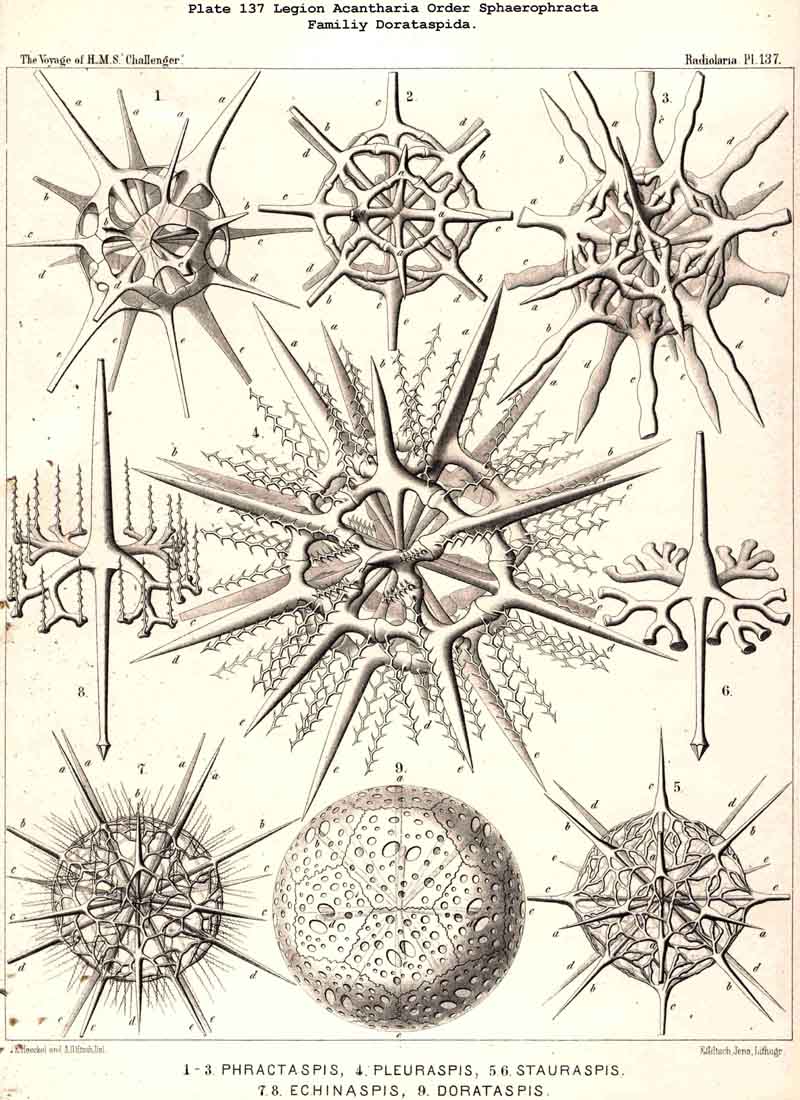
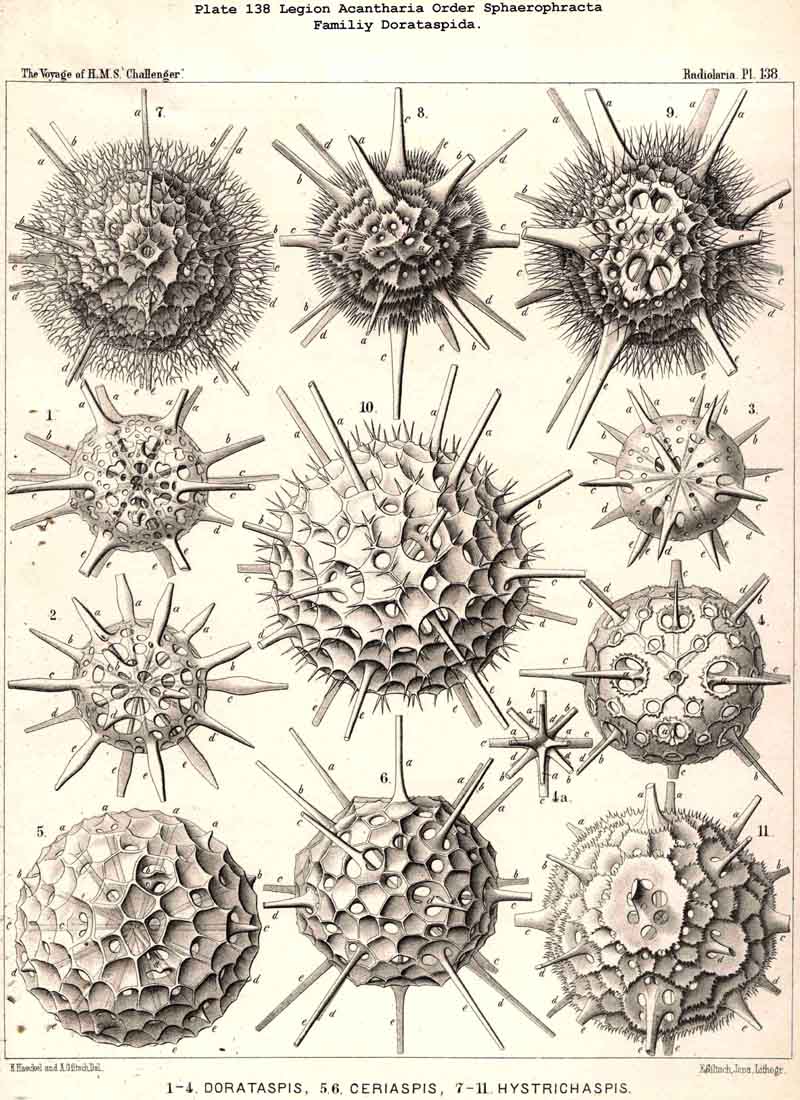
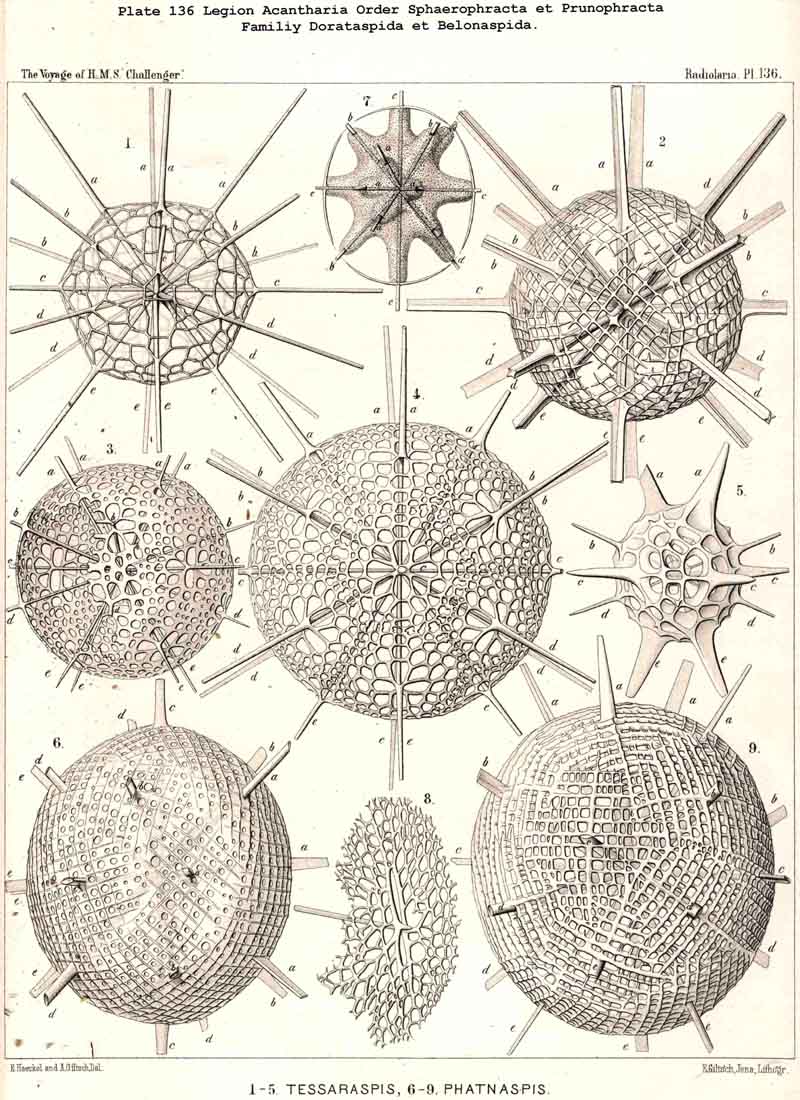
176. Ascent of the Dorataspida.-The group Cladophracta, or those ACANTHARIA whose lattice-shell arises by the union of transverse processes of the twenty radial spines, includes five different families, whose stem-group is the family Dorataspida, with a simple spherical lattice-shell. This family itself is, however, diphyletic in origin, being composed of two essentially and originally different subfamihies-Diporaspida and Tessaraspida (p. 803). The Diporaspida (p. 808, Pls. 137, 138) have been developed from the Phractacanthida, and as each radial spine of the latter bears two opposite apophyses, so the lattice-shell of the former has forty primary aspinab pores (two on the base of each spine). On the other hand, the Tessaraspida (p. 830, Pls. 135, 136) have been developed from the Stauracanthida, and as each radial spine of the latter bears four crossed apophyses, so the lattice-shell of the former has eighty primary aspinal pores (four at the base of each spine).
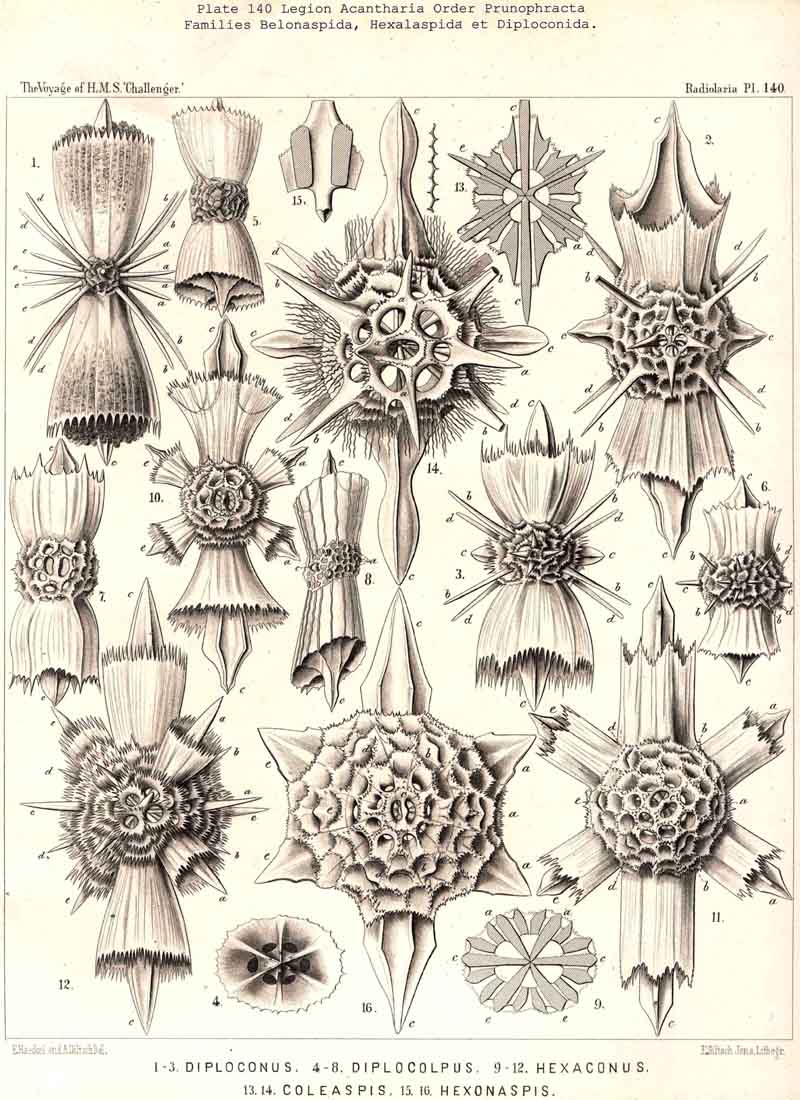
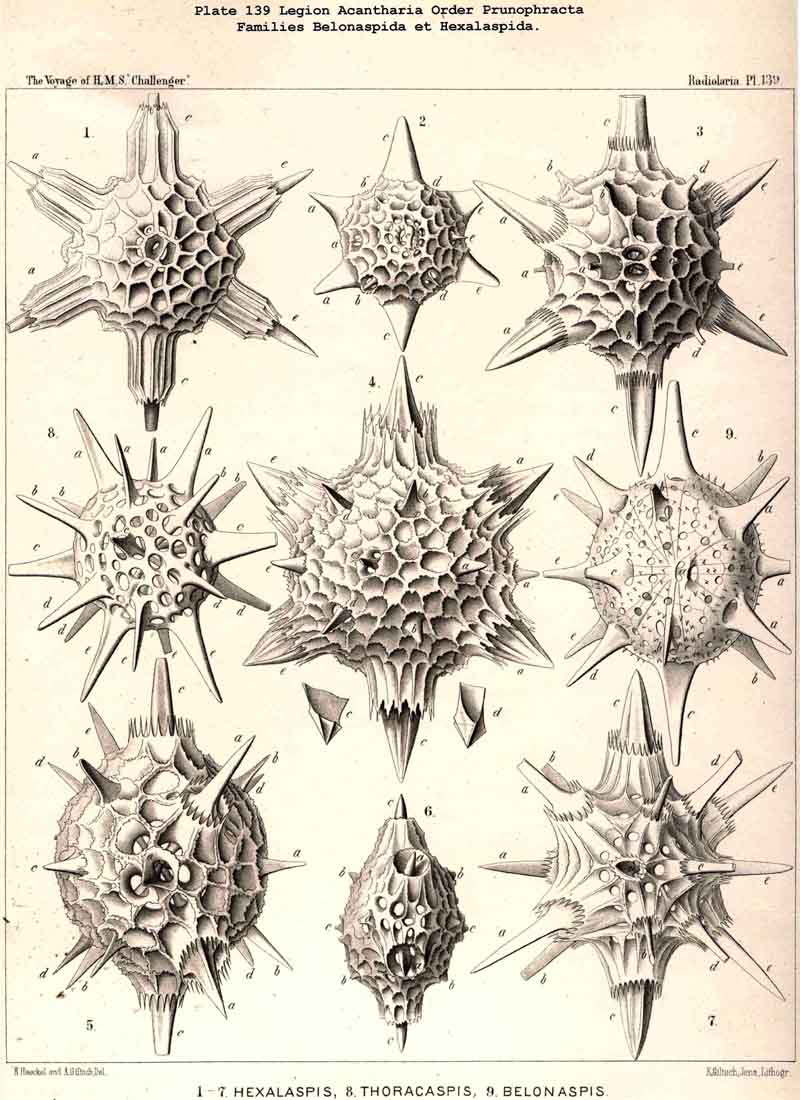
177. Descent of the. Diporaspida.-Whilst the Tessaraspida (§ 176) have given rise to no new groups which could take rank as independent families, no less than four separate families of ACANTHARIA have arisen from the Diporaspida. The Phractopeltida (Pl. 133, figs. 1-6) are distinguished from all other ACANTHARIA by the possession of two concentric. spherical lattice-shells, and have probably been developed from the Diporaspida in the same way as the Dyosphaerida from the Monosphaerida among the Sphaeroidea ; in that case the smaller inner lattice-sphere (medullary shell) would be the primary, and the larger outer sphere (cortical shell) the secondary ; this latter shows forty primary aspinal pores like those of the Diporaspida. The possibility is not excluded, however, that the small inner lattice-sphere of the Phractopeltida is a secondary product. The three remaining families, which must be regarded as descendants of the Diporaspida, form together a single phylogenetic series, and are separated from the primitive group mainly by the fact that the original spherical form of the lattice-shell has been modified into one distinguished by an elongated equatorial axis (the hydrotomical axis) ; hence the Prunophracta (pp. 794-859). The ellipsoidal Belonaspida have arisen directly by hypertrophy of the two opposite equatorial spines of this hydrotomical axis (p. 859, Pl. 136, figs. 6-9 ; Pl. 139, figs. 8, 9 ; perhaps they have also arisen directly from the Amphilonchida). In the lentelliptical Hexalaspida (Pl. 139) all six spines which lie in the hydrotomical meridian plane (two equatorial and four polar) are very strongly developed, the remaining fourteen being rudimentary. Finally, in the Diploconida the two conical sheaths of the two opposite hydrotomical equatorial spines are so predominant that they take the chief part in the formation of the hour-glass-shaped shell.
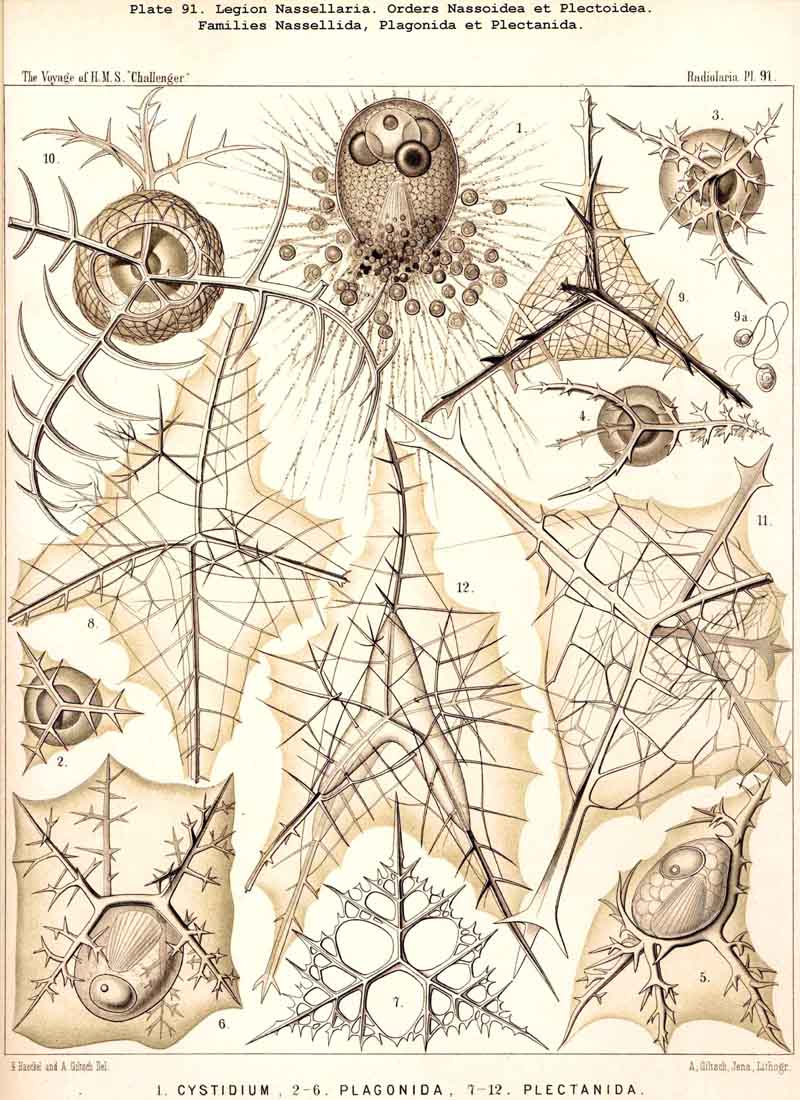
178. Phylogeny of the Nassellaria.- The legion NASSELLARIA or MONOPYLEA is so clearly characterised by the peculiar porochora, which closes the osculum at the oral pole of the monaxon central capsule, and by the podoconus connected with it, that there can be no doubt that phylogenetically it represents an independent stem (§ 8). This stem is only connected at its base by means of Cystidium and Nassella with Actissa and Thalassicolla, the stem-forms of the SPUMELLARIA. This stem is monophyletic, inasmuch as all its members may be derived without violence from the skeletonless Nassellida (Nassella, Cystidium, p. 896, Pl. 91, fig. 1).
179. Origin of the Nassellaria.-The Nassellida (p. 896), which may naturally be considered as the common stem-group of the NASSELLARIA, are most nearly related among other Radiolaria to the Thalassicollida, and in both these skeletonless families the simplest forms, Cystidium and Actissa correspond ; on the other hand, those which have arisen from them by the formation of alveoles in the calymma (Nassella and Thalassicolla) also correspond. The origin of the simplest Nassellida from these primitive Thalassicollida may be explained by supposing that the numerous (formerly evenly distributed) pores of the capsule membrane became obliterated in the upper (apical) half of the central capsule, whilst in the lower (basal) half they became correspondingly more strongly developed ; hence the porochora was formed at the oral pole of the vertical main axis, and a differentiation of the endoplasm proceeding from this gave rise to the characteristic podoconus. Both these organs still at present exhibit very various degrees of progressive development.
180. Hypothetical Genealogical Tree of the Nassellaria.
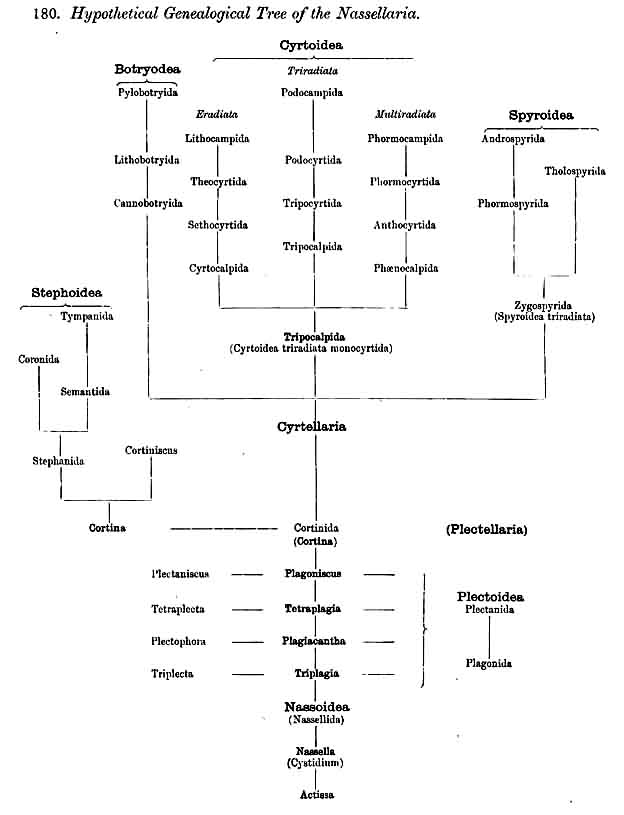 |
181. Plectellaria and Cyrtellaria. -The extensive legion NASSELLARIA far surpasses the other three legions in the endless variety of its skeletal structures, and owing to the complicated relationships of its numerous families presents no lack of difficult phylogenetic problems. All NASSELLARIA may be divided first into two main groups or sublegions, Plectellaria and Cyrtellaria ; the latter having a complete lattice-shell, the former not. Probably the Cyrtellaria. have been polyphyletically developed from several different groups of Plectellaria. These groups are, however, connected in such manifold ways that a monophyletic origin of all the NASSELLARIAN skeletons from one original element is possible. Such a primitive element may have been furnished by any one of three different skeletal parts, the sagittal ring, the basal tripod, and the latticed cephalis (compare pp. 891-895, and Bütschli, L. N. 40, 41).
182. Phylogenetic Skeletal Elements of the Nassellaria.-The multiform skeleton of the NASSELLARIA may be referred in different ways to one of the three above-mentioned structural elements. Each of these (p. 891) may by itself form the skeleton ; the sagittal ring in the simplest Stephoidea (Archicircus, Lithocircus), the basal tripod in the simplest Plectoidea (Triplagia, Plagiacantha), the latticed cephalis in the simplest Cyrtoidea (Cyrtocalpis, Archicapsa). In the great majority of the NASSELLARIA, however, two of these elements, or even all three, are found combined. In most Cyrtellaria, more especially, both the sagittal ring and the basal tripod may be recognised in the lattice-shell, though often only in slight rudiments or scarcely perceptible traces. In the Plectellaria also (which possess no latticed cephahis) there are individual genera with complete development both of the sagittal ring and basal tripod; this important combination is especially well represented in the Cortinida (Cortina, Cortiniscus, Stephanium, Stephaniscus, Tripocoronis, &c.). The greatest difficulty as regards the phylogeny of the NASSELLARIA lies in the fact that the most various combinations of the three elements are presented by closely related or very similar forms. If, in spite of this, a monophyletic hypothesis as to the origin of the NASSELLARIA seems essential all sides of the three possible hypotheses must receive full consideration and critical comparison (§§ 183-191).
183. Ascent of the Nassellaria from the Plectoidea. -The monophyletic hypothesis (No. 2, p. 893) which regards the basal tripod as the common origin of the skeleton of all NASSELLARIA, starts from the simplest forms of the Plectoidea (Triplagia, Plagoniscus, Triplecta, Plectaniscus, &c., Pl. 91). All Plectoidea may be immediately derived as diverging twigs of these, as well as all triradial and multiradial forms of Cyrtoidea and Spyroidea ; for in all these cases the distinctive triradial (or the derived multiradial) form of skeleton appears directly derivable from the simple basal tripod of the former. The same is perhaps also true of many Botryodea Furthermore, certain important forms of Stephoidea (Cortina, Cortiniscus, Stephanium, Stephaniscus, &c.), which have a characteristic combination of the sagittal ring and basal tripod, may be immediately derived from such forms of Plectoidea as Plagoniscus cortinaris, Plagiocarpa procortina, Plectaniscus cortiniscus, &c. On the contrary, those Stephoidea and Cyrtoidea in which the basal tripod is wanting can only be derived from the Plectoidea by the assumption that this structure has disappeared in consequence of phylogenetic degeneration. The monophyletic derivation of the NASSELLARIA from the Plectoidea has more internal probability than that from the Stephoidea, since it is easier to suppose that the Cortinida (Cortina, Stephanium, &c.) have been derived from the Plectoidea (Plagoniscus, Plagiocarpa) than the converse. This view is the basis of the hypothetical tree shown in § 180.
184. Ascent of the Nassellaria from the Stephoidea.-The monophyletic hypothesis (No. 1, p. 893) which regards the primary sagittal ring as the common starting point of the skeleton in all NASSELLARIA, starts from the simplest forms of Stephoidea (Archicircus, Lithocircus, &c., Pl. 81). All Stephoidea and Spyroidea may be immediately derived from these, as also the majority of the Cyrtoidea and probably of the Botryodea. Those numerous forms of the last two groups, however, which possess no trace of a sagittal ring, can only be derived from the former by the supposition that the latter has completely disappeared in consequence of gradual phylogenetic degeneration. The same holds true also of the Plectoidea, although certain forms (e.g., Piagiocarpa procortina, Pl. 91, fig. 5 ; Plectaniscus cortiniscus, Pl. 91, fig. 9) appear to indicate the commencing formation of the sagittal ring by the concrescence of two branches, which approach each other from the upper part of the apical rod and the ventral part of the basal rod. In any case, it is a fact of great phylogenetic significance, that the primary sagittal ring in the cephahis of the Cyrtoidea shows all conceivable stages of degeneration (compare Bütschhi, L. N. 40, 41, as well as the general account of and critical comparison of the NASSELLARIA, pp. 889-895, &c.).
185. Ascent of the Nassellaria from the Cyrtoidea.-The monophyletic hypothesis (No. 3, p. 894) which regards the latticed cephalis as the common point of origin of all the skeletons of the NASSELLARIA, starts from the simplest forms of the Cyrtoidea, that is, from the Cyrtocalpida or eradial Monocyrtida (Archicorida, Archicapsida, Pls. 51, 52, 98). All Cyrtoidea and Botryodea may be regarded as divergent forms of these monothalamous Cyrtoidea ; the polythalamous simply by the addition of fresh joints at the basal pole, the triradiate and multiradiate by the development of three or more apophyses. The origin of the sagittal ring (which presents every stage of development and degeneration in the Cyrtoidea) may be regarded as a mechanical thickening of the latticed plate in the sagittal circumference of the cephahis. By stronger development of this ring and coincident sagittal constriction of the cephahis the order Spyroidea may be derived from the Cyrtoidea. On the other hand, the Plectellaria, which possess no cephalis, and indeed no complete lattice-shell what ever, may be derived from the Monocyrtida by the assumption of a. degeneration of this structure ; the sagittal ring having been preserved in the Stephoidea, and the tripod of the Tripocalpida in the Plectoidea. Although such a monophyletic derivation of the NASSELLARIA from the Cyrtocalpida is possible, and though here, too, the Cortinida play an important part as connecting links, this hypothesis bias less internal probability than that of the derivation from the Stephoidea (§ 184) or Plectoidea (§ 183).
186. Genealogical Tree of the Plectoidea.-The order Plectoidea includes those NASSELLARIA whose rudimentary skeleton does not contain the characteristic sagittal ring of the Stephoidea, but consists of several (at least three) radial spines, which proceed from a point in the centre of the porochora. The branches of these radial spines remain free in the Phagonida, whilst in the Plectanida they unite with each other to form a loose meshwork (not, however, a complete lattice-shell). The number and arrange ment of the radial spines, which serve for generic distinctions, are the same in both families, so that each genus of the Plectanida has arisen from a corresponding genus of the Plagonida. The simplest Plagonida, which possess a basal tripod (Triplagia or Plagiacantha with three rays, Tetraplagia with four rays) are probably to be regarded as forming the common origin of the whole order. These agree with certain three- and four-rayed skeletal pieces of the Beloidea (Thalassosphaerida and Sphaerozoida) ; and also the four and six-rayed twinned pieces of the latter (spicula bigemina and trigemina) repeat in the same fashion the skeleton of the former (Plagonidium, Plagonium). This similarity, however, is a mere analogy and possesses no phylogenetic significance. On the other hand, certain Plagonida (Plagoniscus, Plagiocarpa), and the corresponding genera of Plectanida (Plectaniscus, Periplecta) seem to have important phylogenetic relations to certain Stephoidea (Cortina, Cortiniscus, &c.) ; the sagittal ring of the latter having perhaps arisen by the vertical apical spine of the former having been connected with their horizontal basal rod by two ventral apophyses growing out opposite to each other (compare pp. 902, 914, Plagiocarpa procortina, Pl. 91, fig. 5). In this case the Plectanida would belong to the simplest stem-forms of the NASSELLARIA.
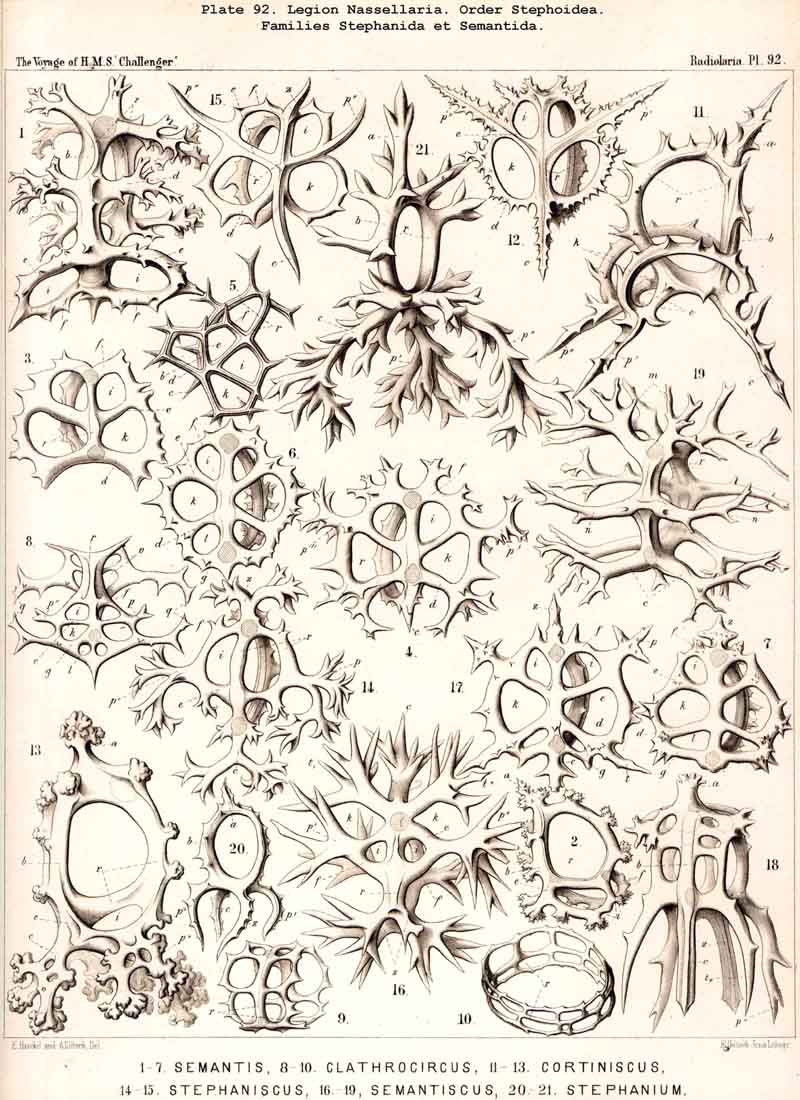
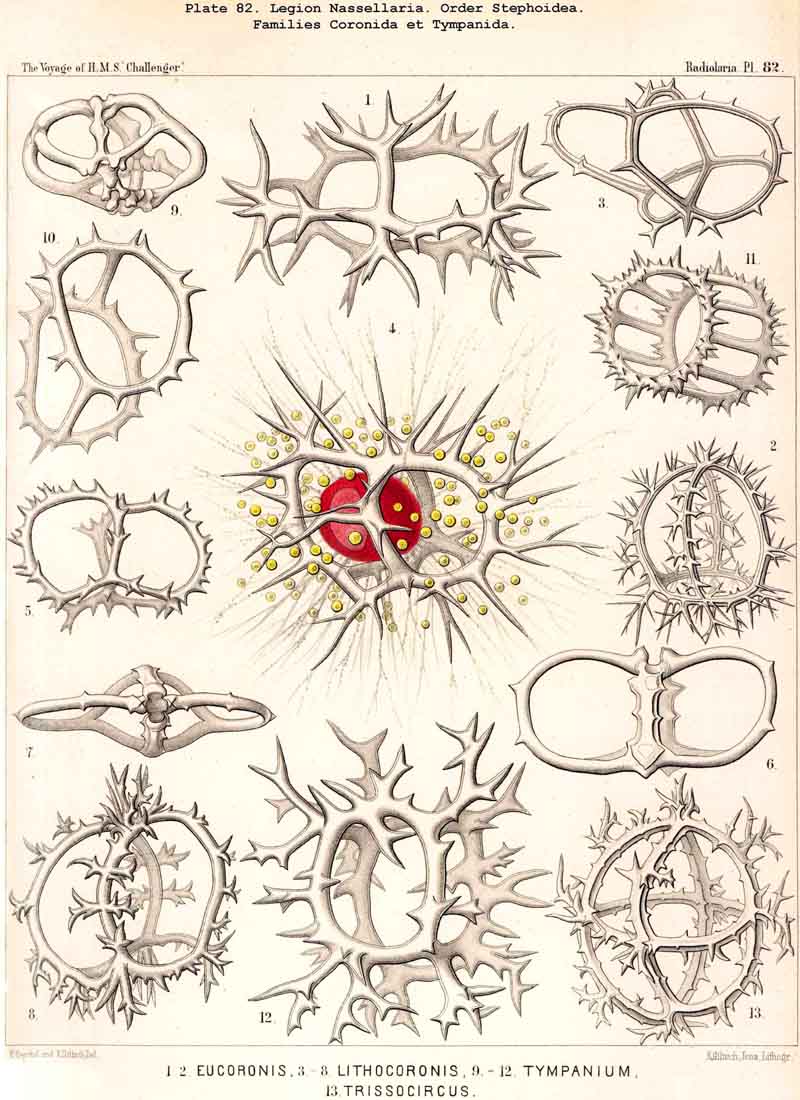
187. Genealogical Tree of the Stephoidea.-The order Stephoidea includes all those NASSELLARIA whose skeleton does not form a complete lattice-shell, but consists of one or more rings, and often of a loose meshwork which arises by the union of branches of the rings. A vertical sagittal ring is constantly present, embracing the central capsule in the median sagittal plane, and forming at its basal pole various processes, the starting point for other skoletal forms. The most important of these is the tripodal Cortina (p. 950, § 182). The Stephanida are the most archaic family among the Stephoidea (p. 937, Pl. 81), perhaps indeed among all the NASSELLARIA (§ 184); in them the sagittal ring and its processes alone constitute the skeleton ; secondary rings and meshes are wanting. Two diverging families, the Semantida and Coronida, have been developed from the Stephanida, and from one of them the family Tympanida has arisen.
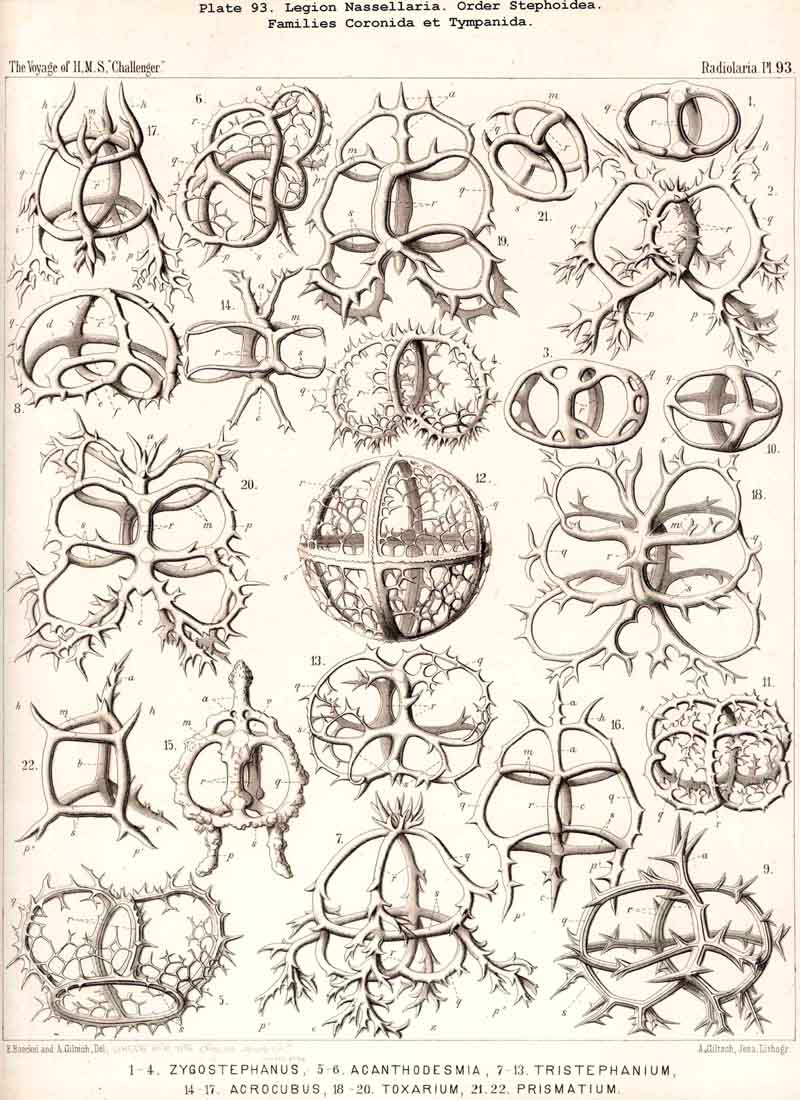
The Semantida (p. 953, Pl. 92) develop a horizontal basal ring at the oral side of the vertical sagittal ring ; the basal meshes or lattice gates, which remain between the former and the latter, are the important cortinar pores (one pair jugular, one pair cardinal, p. 954); they usually appear inherited in the cortinar septum of the Cyrtellaria. In the Coronida (p. 967, Pls. 82, 94) a second vertical ring (the frontal ring) appears in addition to the sagittal ring ; it lies in the frontal plane at right angles to the latter. Finally the Tympanida (p. 987, Pls. 93, 94) have probably arisen from the Semantida by the formation of a second horizontal ring (mitral ring) parallel to the basal and attached to the upper portion of the sagittal ring.
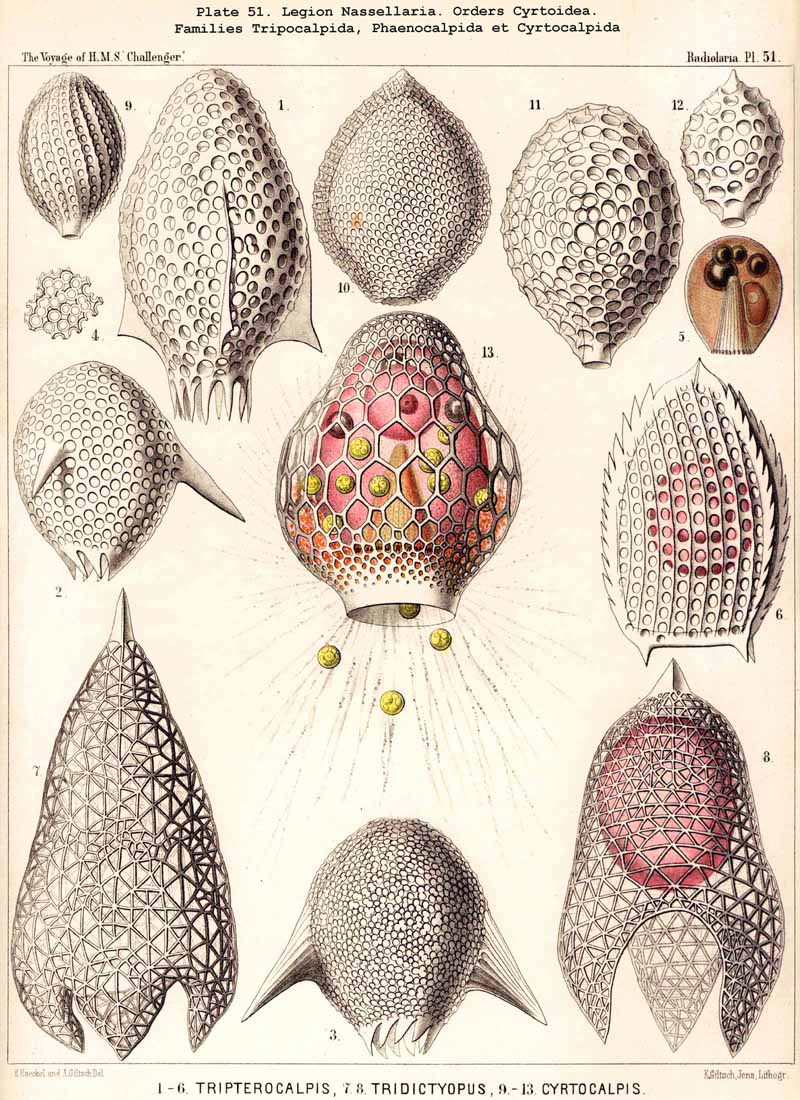
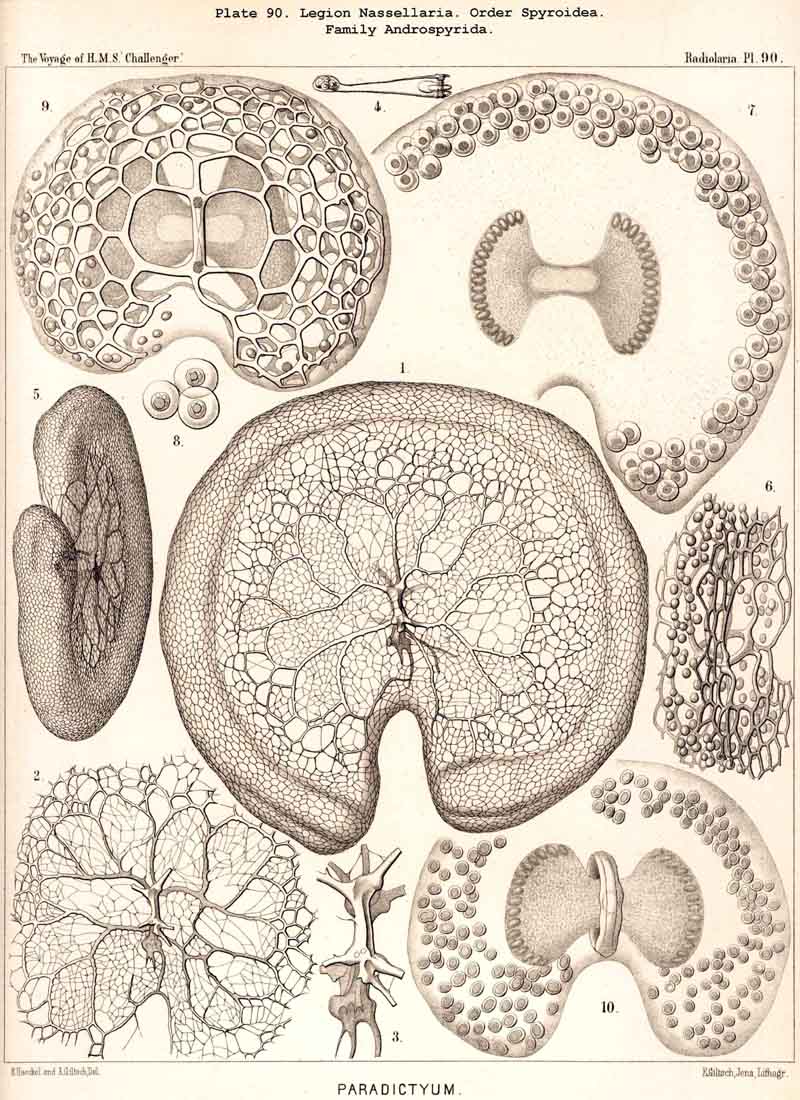
188. Genealogical Tree of the Spyroidea.-The extensive order Spyroidea is of especial interest in connection with the phylogeny of the NASSELLARIA, since all its members show two well-developed skeletal elements in combination, the sagittal ring of the Stephoidea and the latticed cephahis of the Cyrtoidea; the majority possess also the basal tripod of the Plectoidea (or a radial skeleton derived from it). Hence there is a possibility of deriving the stem-forms of the Spyroidea from each of these three groups. The four families of this order exhibit similar relationships to those of the four families of Cyrtoidea ; the common stem-group is the family Zygospyrida ; from this the Tholospyrida have arisen by the development of a galea on the apical pole, the Phormospyrida by the addition of a thorax on the basal pole. The Androspyrida may be derived either from the Tholospyrida by the formation of a basal thorax, or from the Phormospyrida by the development of an apical galea. Some groups, however, such as the peculiar Nephrospyrida (Pl. 90) have probably been developed directly from the Stephoidea.
189. Genealogical Tree of the Botryodea. -The peculiar order Botryodea (p. 1103), which is both difficult to investigate and insufficiently known, presents great phylogenetic difficulties both as to its ascent and descent. Probably the different genera of this order have been polyphyletically developed from different groups of Cyrtoidea (perhaps also to some extent of Spyroidea) by the formation of lobes in the cephalis. The three families of Botryodea are related to each other in the same way as are the three first families of the Cyrtoidea. From the single-jointed Cannobotryida (corresponding to the Monocyrtida), the two-jointed Lithobotryida (corresponding to the Dicyrtida), may be derived by the development of a basal thorax, and from the latter the three-jointed Pylobotryida (like the Tricyrtida) by the addition of an abdomen. In the last two families the forms with an open basal mouth (Botryopylida and Botryocyrtida) are to be regarded as primitive ; the Botryocellida and Botryocampida have arisen by the closure of this mouth with a basal lattice-plate.
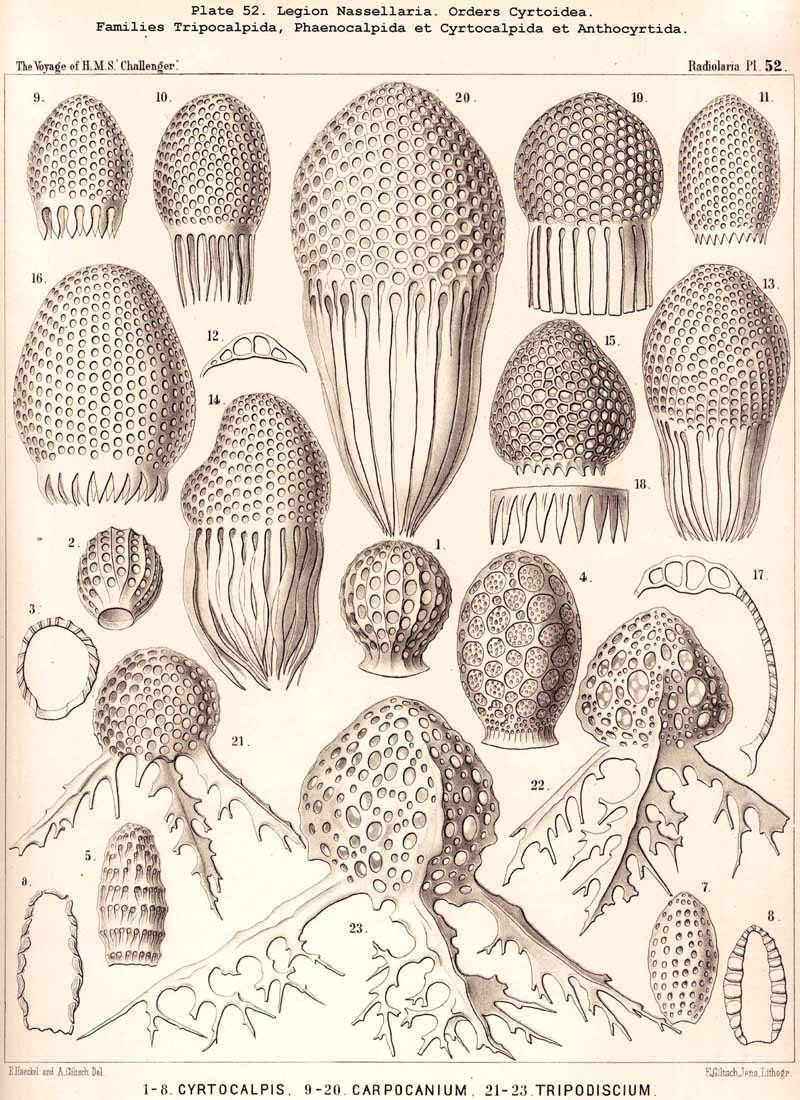
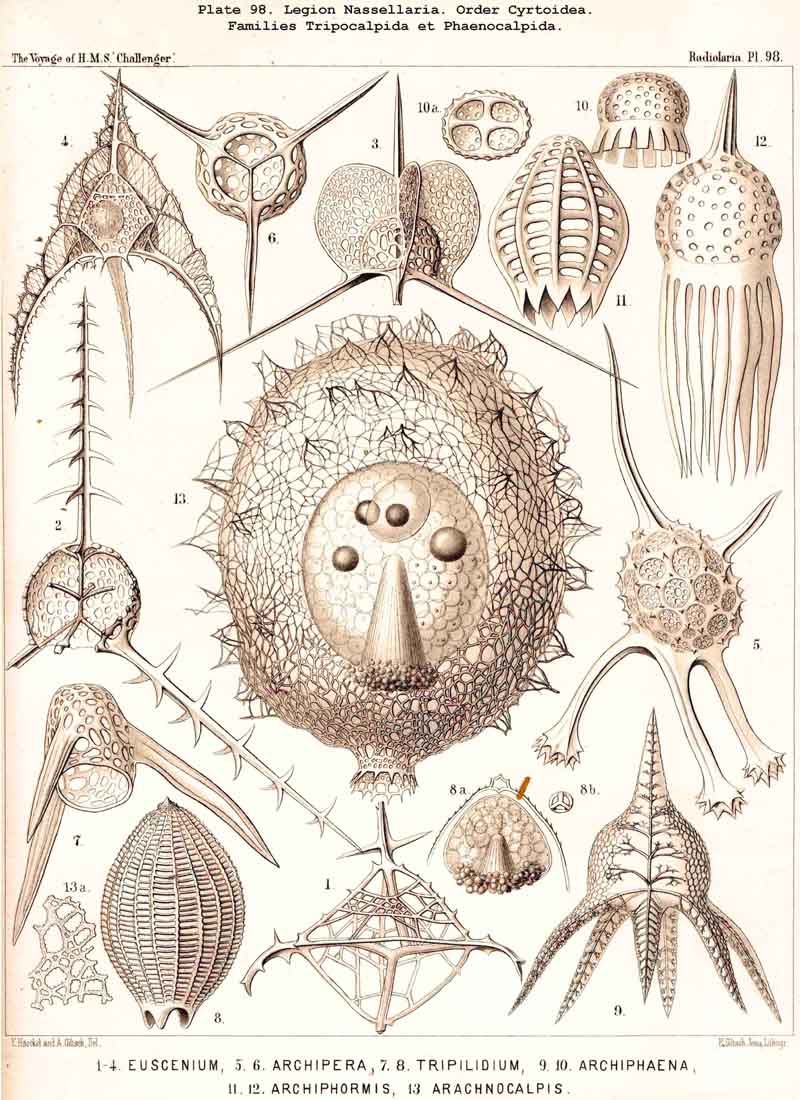
190. Genealogical Tree of the Cyrtoidea..-The multiform and extensive group Cyrtoidea presents the greatest difficulties to be found in the phylogeny of the NASSELLARIA, because their morphological relations are most complicated, and because similar forms very often appear to be of quite different origin. The great majority of the Cyrtoidea show more or less clearly a combination of the three structural elements ; sagittal ring, basal tripod, and latticed cephahis (p. 891). There are also, however, numerous Cyrtoidea, whose skeleton no longer shows any trace of the sagittal ring. Many of these show as the basis of the skeleton a strong basal tripod with an apical spine, around which the cephalis has obviously been secondarily developed, e.g., the remarkable Euscenida (p. 1146, Pls. 53, 97) and the interesting Callimitrida (p. 1217, Pls. 63, 64). These may have been derived immediately from the Plectoidea without any relation to the Stephoidea. There are also numerous true Monocyrtida, whose shell consists of a simple latticed cephalis without a trace of the sagittal ring or basal tripod (Cyrtocalpida, Pl. 51, figs. 9-13 ; Pl. 98, fig. 13); these may have been developed directly from the skeletonless Nassellida by the formation of a simple ovoid Gromia-like shell, and may have no relation either to the Stephoidea or Plectoidea. On these grounds, as well as from the complicated relationships of the many smaller groups of Cyrtoidea, it is probable that the whole order has been developed polyphyletically from different divisions of the Plectellaria.
191. Systematic Arrangement of the Cyrtoidea.-Although for the reasons just given no systematic arrangement of the Cyrtoidea can at present, or for a long time in the future, be regarded as other than artificial, yet some general principles of classification for this extensive group can be laid down, which may serve as starting points for some future natural disposition. This is especially true of the relations which in an artificial system (p. 1129) were primarily utilised for the distinction of twelve families and twenty-four subfamilies ; the number of segments in the shell, the number of radial apophyses (and parameres), and the constitution of the basal aperture of the shell.
As regards the number of segments, separated by transverse constrictions, of which the shell is composed, it is dependent upon the secondary addition of new joints at the basal pole of the main axis. Hence all many-jointed Cyrtoidea are to be derived from single-jointed ones, and the four sections thus distinguished (Monocyrtida, Dicyrtida, Tricyrtida, Stichocyrtida) form a phylogenetic series. Very often, however, the primary cephahis disappears owing to retrograde metamorphosis ; and in such cases the single joint of the apparent Monocyrtida is formed of the thorax (e.g., Pls. 52, 54, &c.). As regards the number of radial apophyses, three sections of Cyrtoidea may be distinguished ; the Pilocyrtida with three, the Astrocyrtida with numerous apophyses, and the Corocyrtida with none (p. 1129). The last two may in general be regarded as two divergent branches from the first, for the eradiate Corocyrtida have probably been formed from the triradial Piocyrtida by entire loss of the radial apophyses, whilst on the other hand the multiradiate Astrocyrtida have arisen from them by additions to the three primary apophyses (interpolation ot interradial between the perradial ones). As regards the constitution of the shell-aperture, the Cyrtoidea may be divided into Cyrtaperta and Cyrtoclausa (p. 1129) ; in general the Cyrtoclausa (with latticed shell-aperture) have arisen from the Cyrtaperta (with simple open mouth) ; in many Monocyrtida the converse may be supposed, the simple basal mouth having been formed by degeneration of a basal lattice.
192. Phylogeny of the Phaeodaria.-The legion PHAEODARIA or CANNOPYLEA is so clearly marked off from other Radiolaria by the double membrane of the central capsule and the astropyle at its oral pole, as well as by the extracapsular phaeodium, that it must be regarded phylogenetically as an independent stem (§ 9). This stem is only connected at its root by Phaeodina with the stem-form of the SPUMELLARIA, Actissa. The stem itself is monophyletic, inasmuch as its members may be derived without violence from the skeletonless Phaeodinida (Phaeodina, Phaeocolla). On the other hand, the formation of the skeleton of the PHAEODARIA is undoubtedly polyphyletic, different Phaeodinida having independently commenced the formation of a skeleton, and having carried it out in very different ways.
193. Origin of the Phaeodaria.-The Phaeodinida (p. 1544, Pl. 101), which may naturally be regarded as the common stem-group of the PHAEODARIA, have their nearest relations among other Radiolaria in the Thalassicollida (p. 10); and since this family is to be regarded as the primitive group of all Radiolaria, they may be directly, derived from them phylogenetically. The essential modifications by which the primitive Phaeodinida have arisen from the more archaic Thalassicollida are of three kinds ; (1). the doubling of the membrane of the central capsule ; (2) the reduction of the numerous fine pores in the membrane and the formation of an osculum, and of an astropyle closing it, at the oral pole of the main axis ; (3) the production of an extra capsular phaeodium. This last may, perhaps, be regarded as a unilateral hypertrophy of the voluminous pigment masses which are deposited in the sarcomatrix of certain Thalassicollida. Of the two genera of Phaeodinida hitherto known, probably Phaeodina (Pl. 101, fig. 2) approaches the original stem of the PHAEODARIA more nearby than Phaeocolla (Pl. 101, fig. 1), for the latter exhibits only the large main opening of the central capsule (astropyle), whilst the former possesses also a pair of accessory openings (parapylae). The hypothetical stem-form (Phaeometra) presumably had a larger number of Small parapylae (hike many Circoporida and Tuscarorida), and the astropyle was probably but little differentiated from them.
194. Hypothetical Genealogical Tree of the Phaeodaria .
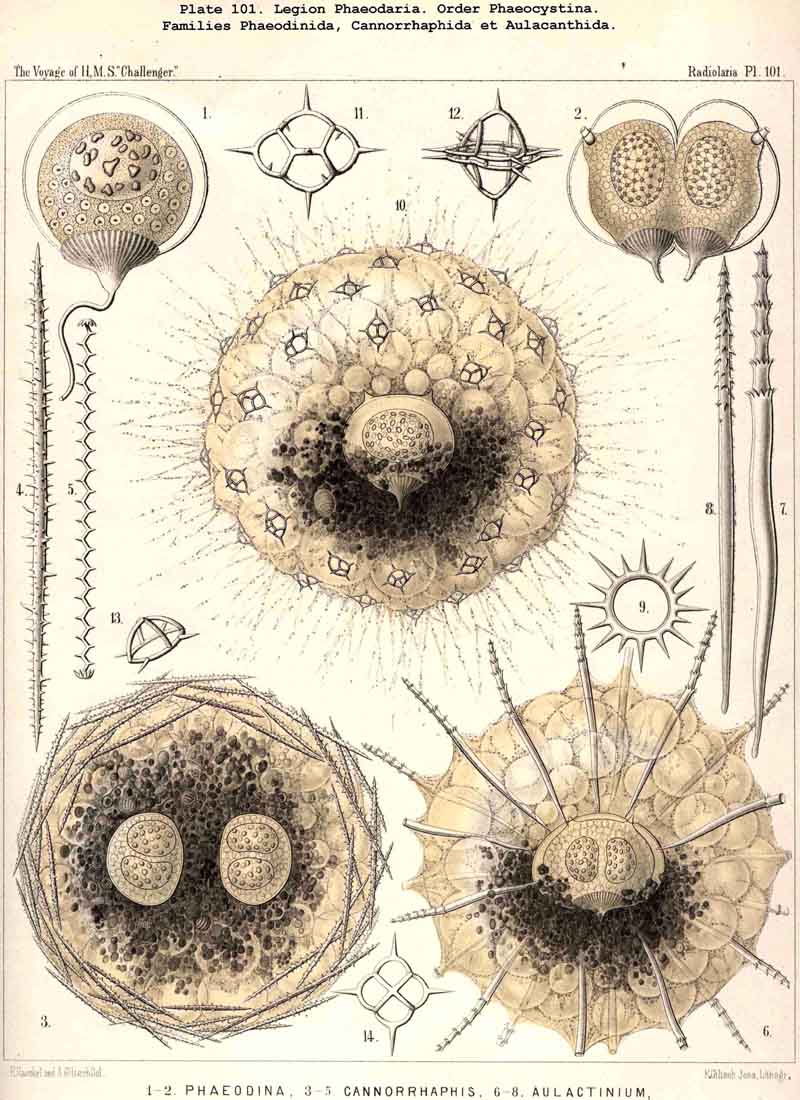
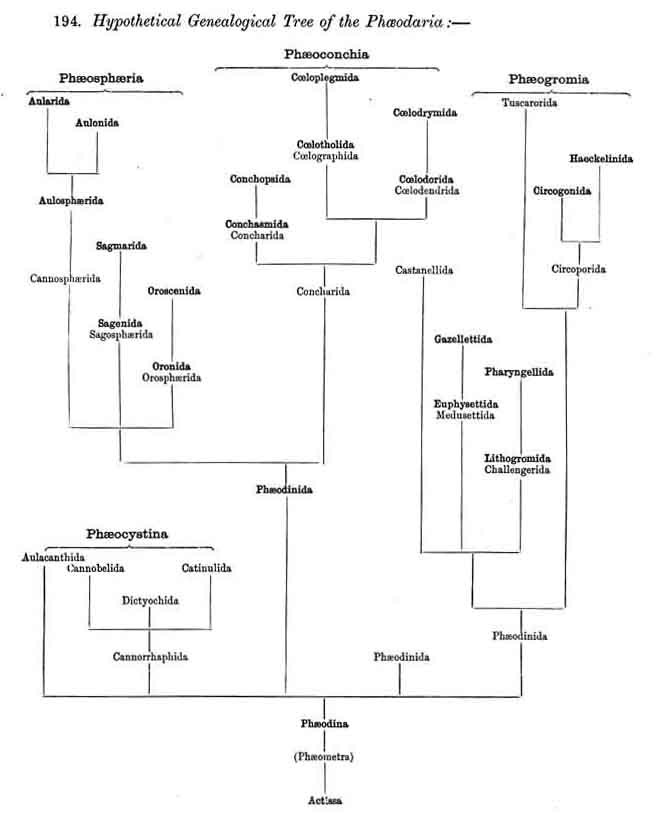 |
195. Phaeocystina and Phaeocoscina.-Whilst the malacoma of all PHAEODARIA possesses the characteristics of the legion, and hence justifies the assumption of a monophyletic origin, the skeleton, on the other hand, shows in the different groups such manifold and fundamental variations that a polyphyletic origin of the latter is indubitable. Different Phaeodinida have commenced the formation of the skeleton independently, and it has progressed in different directions. In the Phaeocystina remained incomplete and led to the formation of various Beloid skeletons, whilst the Phaeocystina developed complete lattice-shells. Both of these divisions too are to be regarded as polyphyletic, since the skeletal forms of the different groups cannot be derived without violence from a common primitive form.
196. Phaeocystina with a Beloid Skeleton.-The order Phaeocystina includes all PHAEODARIA which have no complete lattice-shell ; it contains, firstly, the skeletonless Phaeodinida (the common stem-group of the legion), and secondly, the Phaeacanthida, or PHAEODARIA with a Beloid skeleton (§ 115). The latter are divisible into several very different groups (at least two or three) which are probably. different in origin. The Aulacanthida (Pls. 102-105) form radial tubes which perforate the calymma, their proximal end resting upon the surface of the central capsule, whilst the distal extremity projects freely outwards. The skeleton of the Cannorrhaphida, on the other hand, is composed of many separate portions which are never radially arranged but are either placed tangentially to the surface of the calymma or scattered irregularly in its gelatinous mass. Furthermore, in the three subfamilies of which this family is composed, the individual skeletal portions are so different that they have probably arisen inde pendently of each other ; in the Cannobelida they form cylindrical tangential tubes (Pl. 101, figs. 3-5), in the Catinulida flat basin or cap-like structures (Pl. 117, fig. 8), in the Dictyochida hollow rings, from which small pyramids are developed by unilateral formation of lattice-work (Pl. 101, figs. 9-14 ; Pl. 114, figs. 7-12).
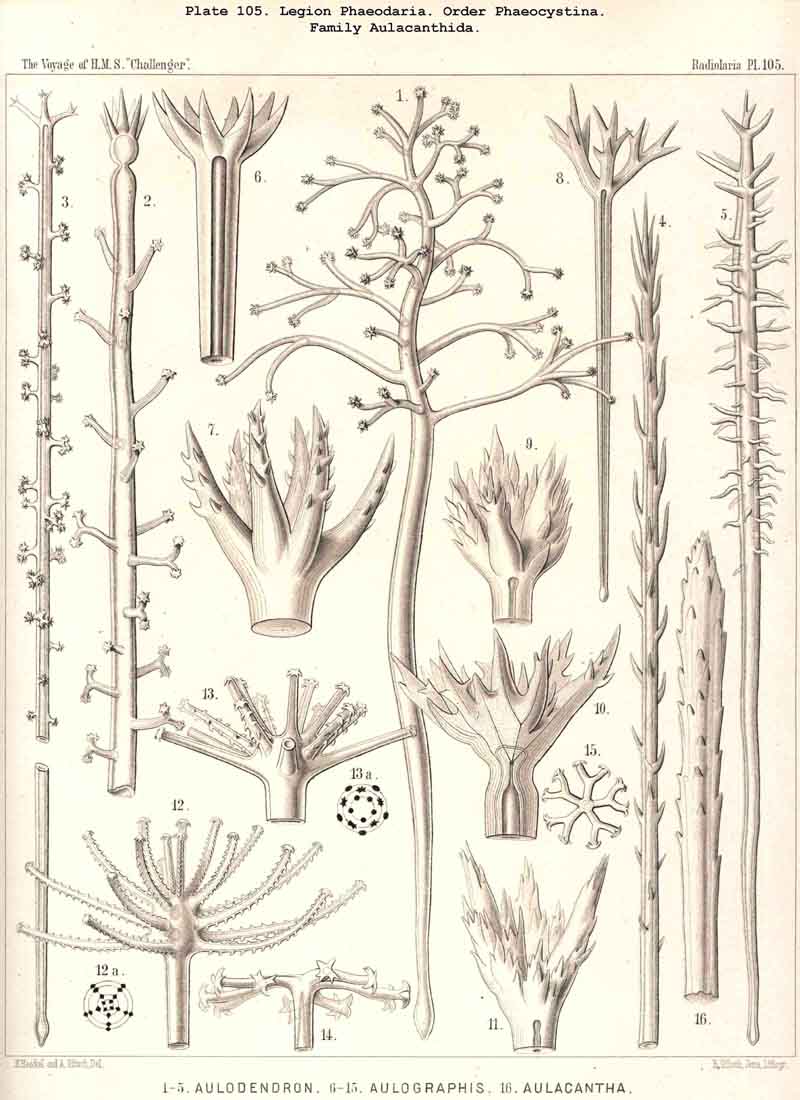
197. Phaeosphaeria with a Sphaeroid Skeleton.-The order Phaeosphaeria includes those PHAEODARIA which possess a spherical (sometimes slightly modified) lattice-shell without the characteristic aperture of the Phaeogromia. They have probably arisen independently of these, though they may have been derived from the Castanellida by loss of the shell-aperture, which was present originally. The four families which we have distinguished among the Phaeosphaeria, are so different in the structure of their lattice-shell that their phylogenetic connection is doubtful. In the Orosphaerida (Pls. 106, 107) and the Sagosphaerida (Pl. 108) the whole lattice-shell consists of a single piece and is unjointed (without astral septa) ; in the former it is very firm and massive, with thick laminated trabeculae and polygonal meshes ; in the latter it is very delicate and brittle, with filiform trabeculae and large triangular meshes. On the other hand, the voluminous shell of the Aulosphaerida (Pls. 109, 110, 111), and of the Cannosphaerida (Pl. 112), is characterised by a very peculiar system of joints ; it is composed of numerous separate cylindrical tubes, which are placed tangentially and united at the nodes by stellate partitions or astral septa. The Cannosphaerida possess further a simple central Cyrtoid shell, connected with the outer jointed shell by hollow radial trabeculae. Since many Aulosphaerida possess rudiments of such centripetal trabeculae it is possible that these latter have been derived from the former by the loss of the central Cyrtoid shell ; the formation of this monaxon shell perhaps indicates descent from the Phaeogromia (Castanellida).
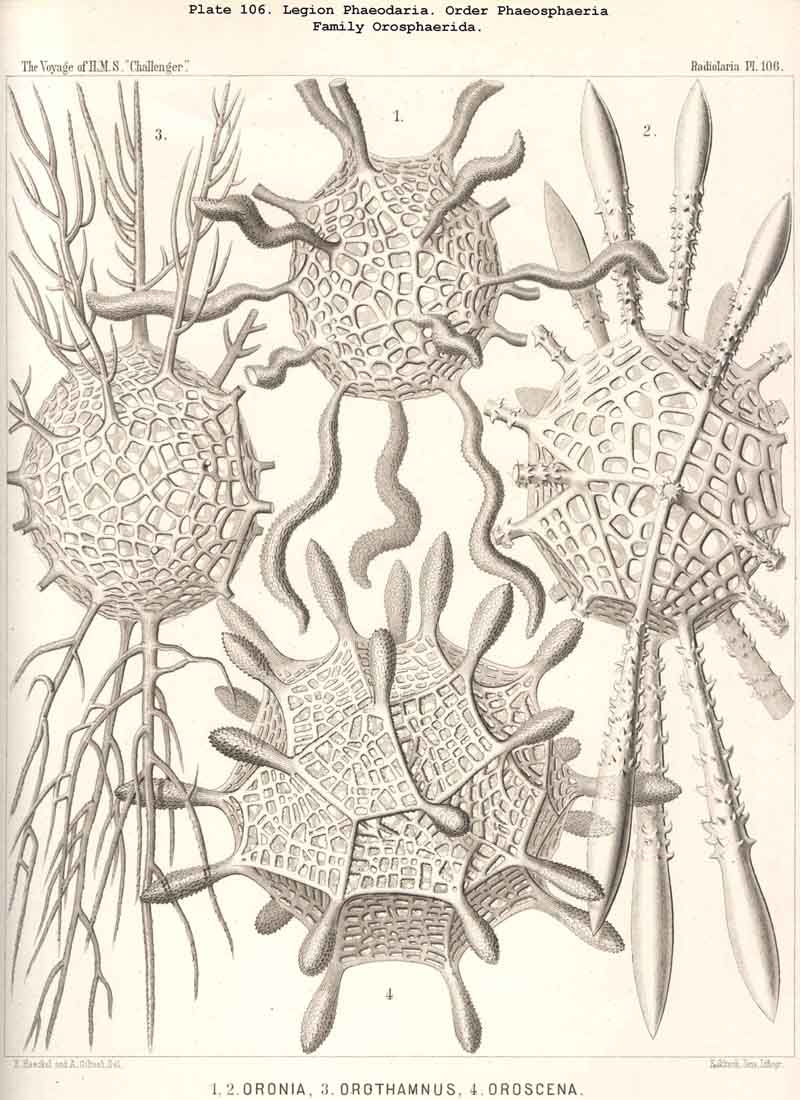
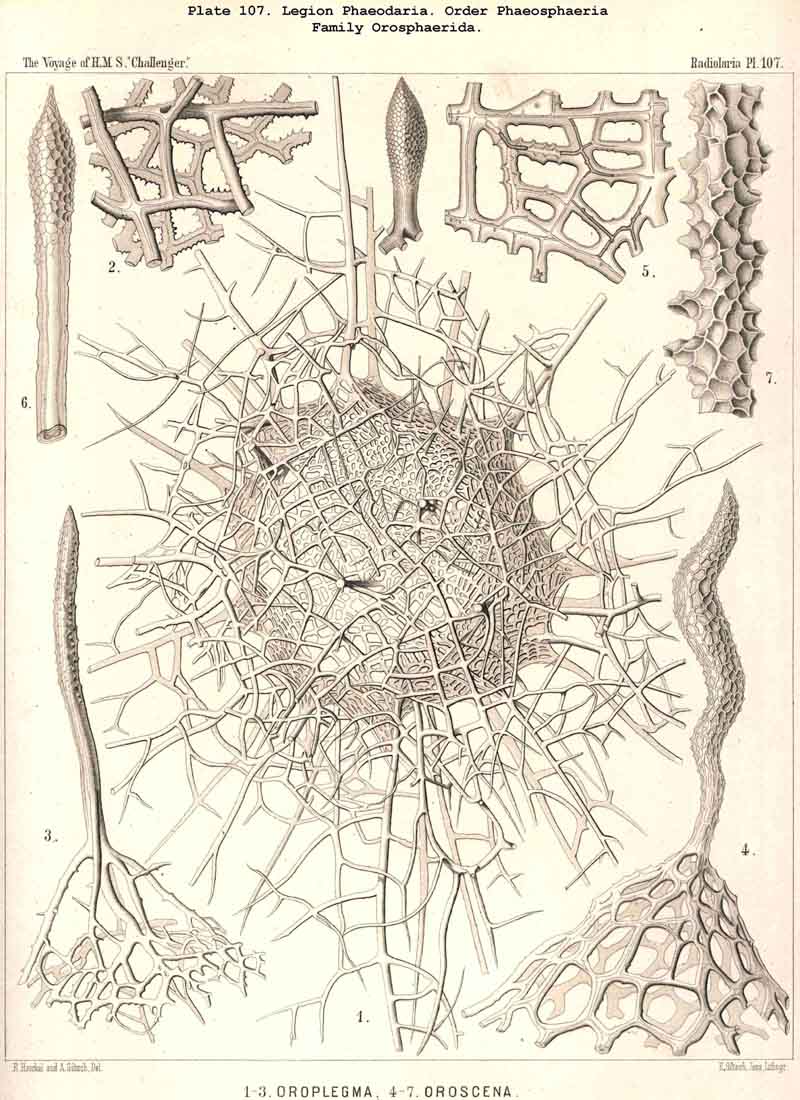
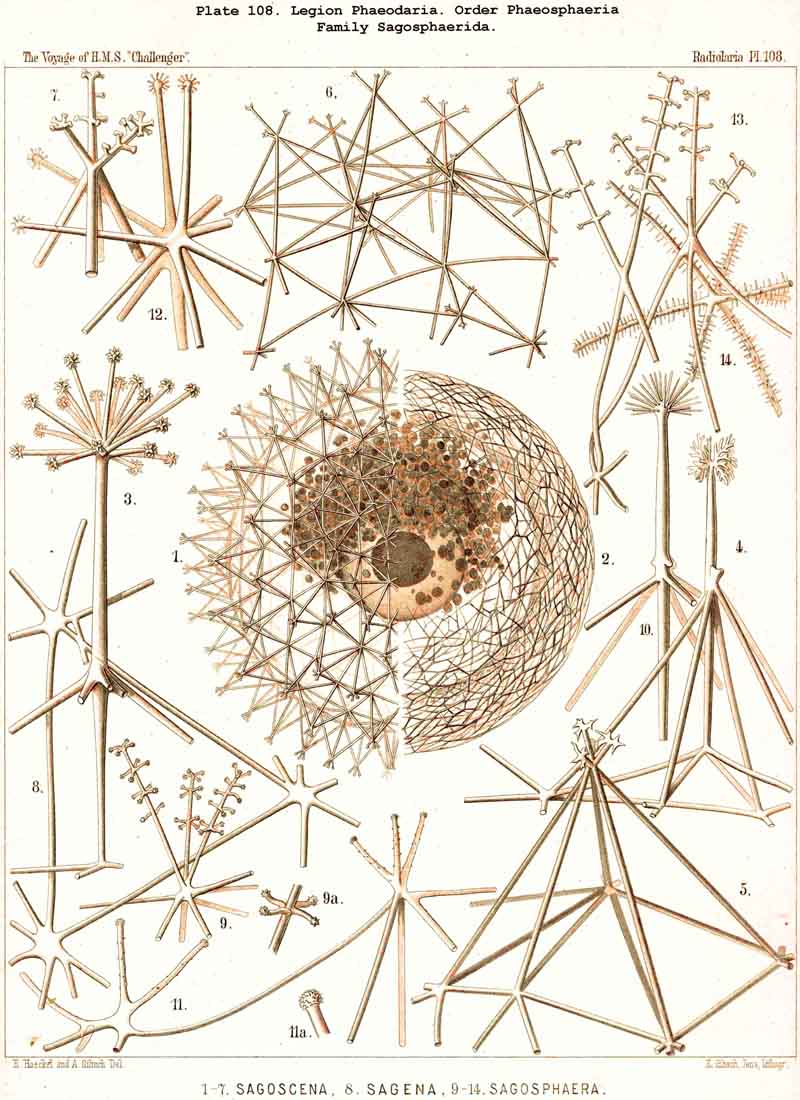
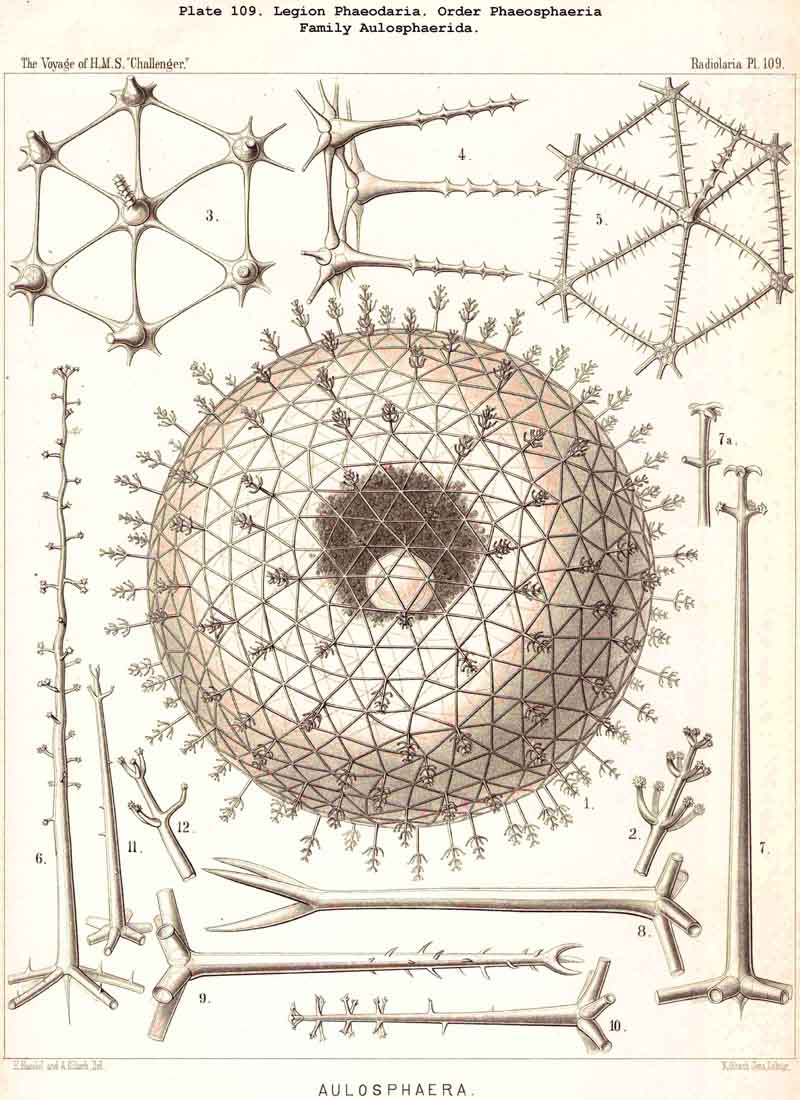
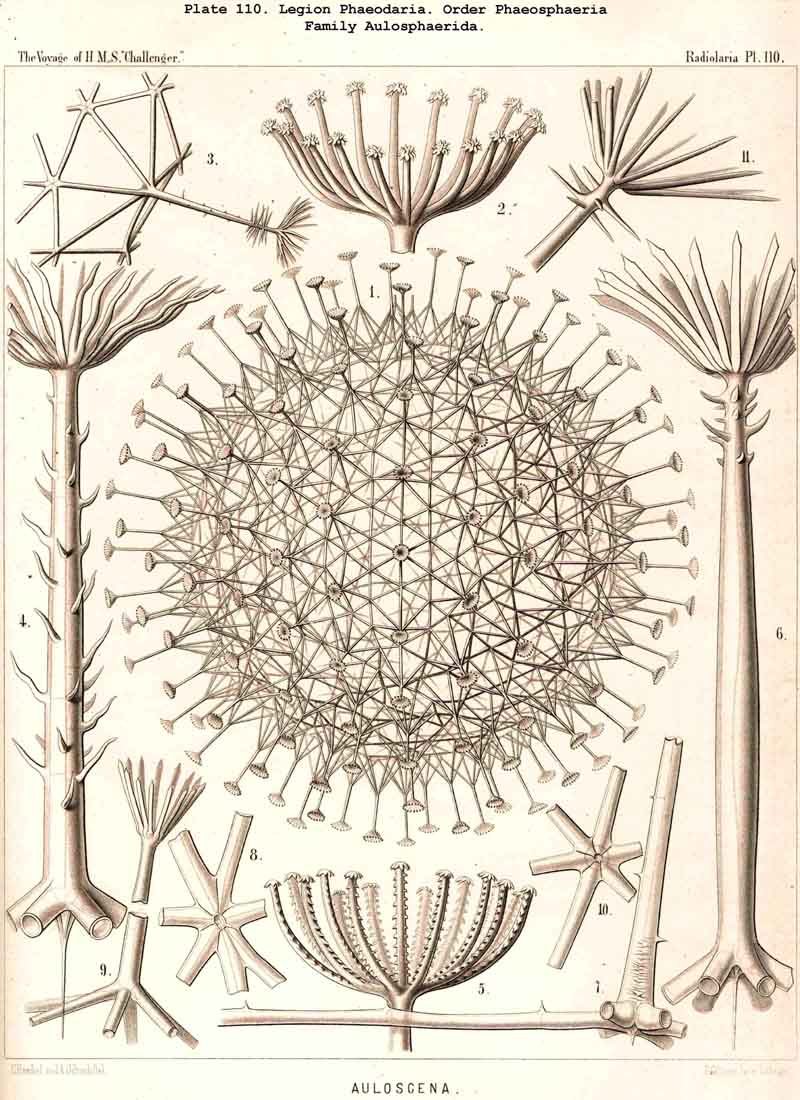
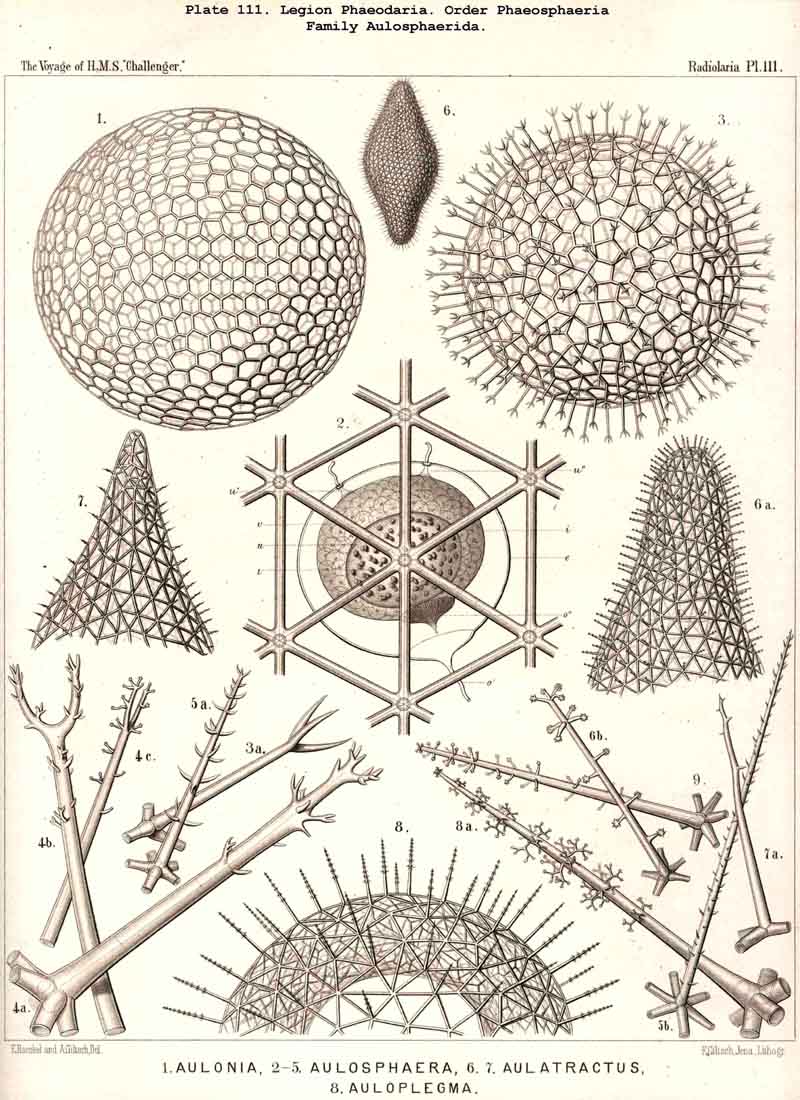
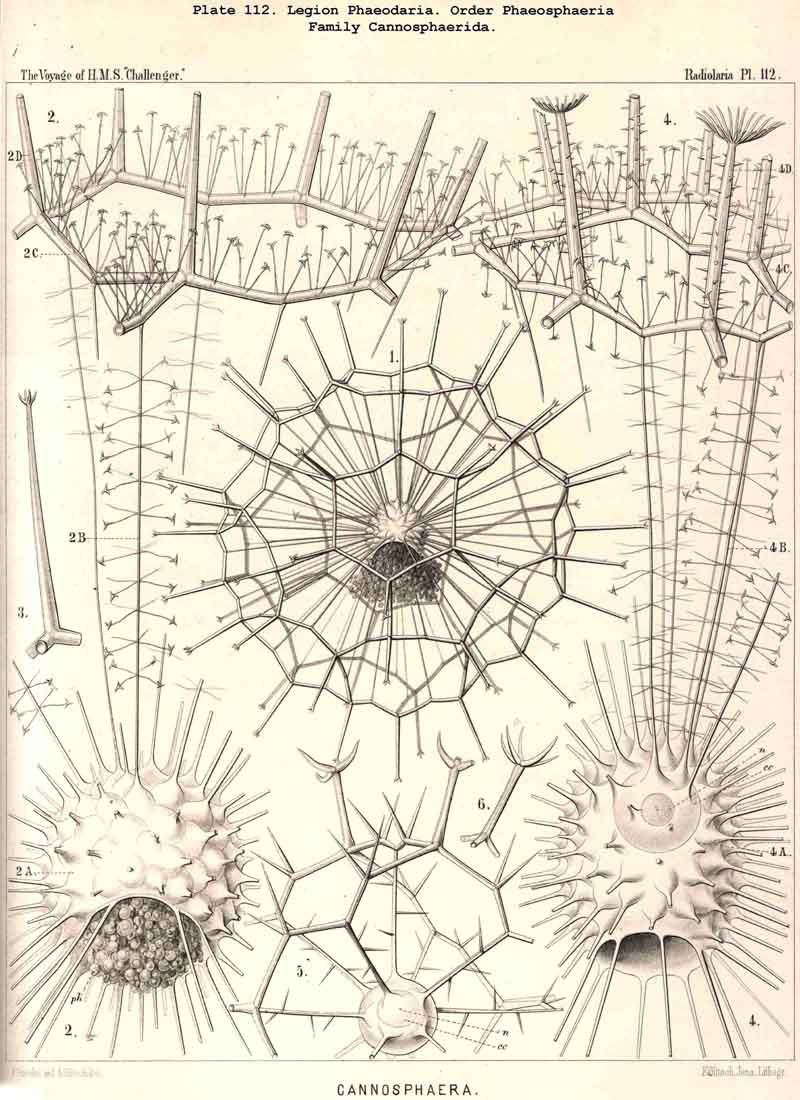
198. Phaeogromia with a Cyrtoid Skeleton.-That order of the PHAEODARIA which we designate Phaeogromia, contains many very different forms, all agreeing in the possession of a Cyrtoid skeleton, or a monaxon lattice-shell, which has a large aperture at one pole of its vertical main axis (§ 123). This Cyrtoid skeleton is sometimes ovoid or conical, sometimes lentiform or helmet-shaped, sometimes polyhedral or almost spherical. Although the principle of its structure is simple and often very like, that of the Monocyrtida among the NASSELLARIA, yet the structure of the wall and of the apophyses is so different in the various groups of the Phaeogromia, that the order is probably polyphyletic, and its Cyrtoid shells have arisen independently of each other. Only in the Castanellida (Pl. 113) does the shell-wall usually consist of simple lattice-work ; in the Challengerida, on the other hand (Pl. 99), it has an extremely fine Diatom-hike structure ; in the Medusettida (Pls. 118-128) a peculiar alveolar structure,. and in the Circoporida (Pls. 114-117) and Tuscarorida (Pl. 100) it possesses a characteristic porcellanous constitution (with tangential spicules in a porous cement-mass) ; in the latter of these groups the surface is smooth, in the former peculiarly tabulate ; the two families have also different stem-forms.
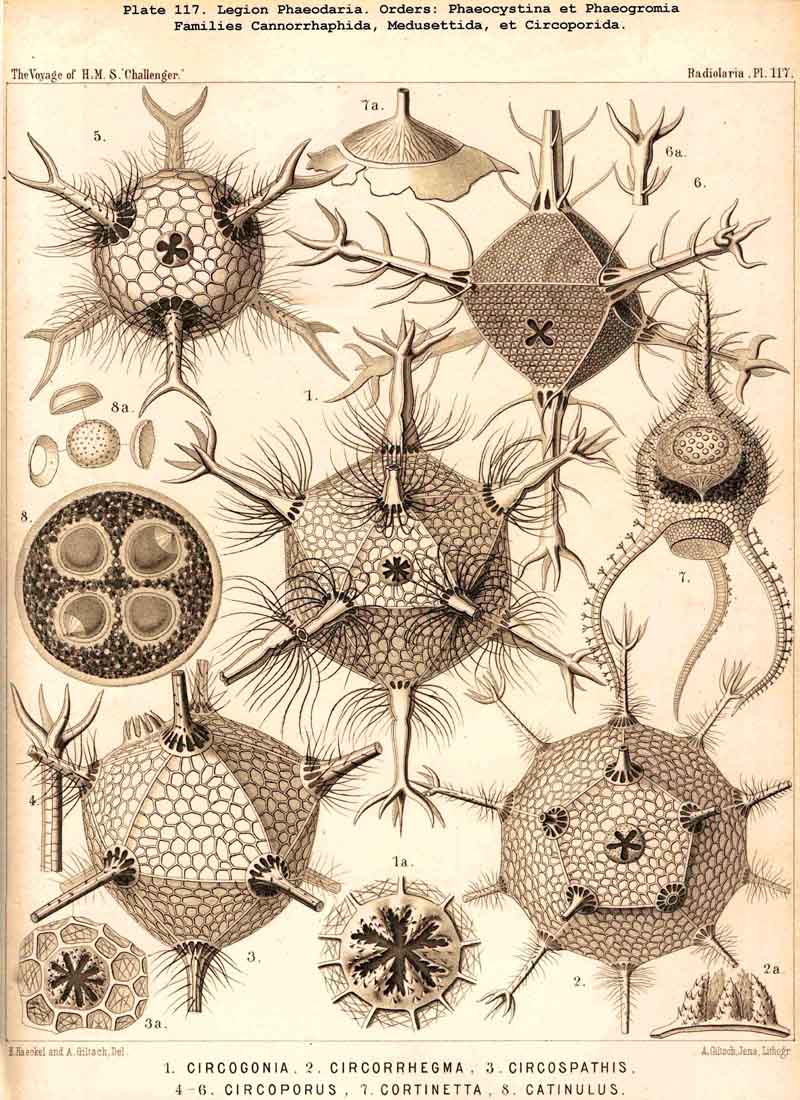
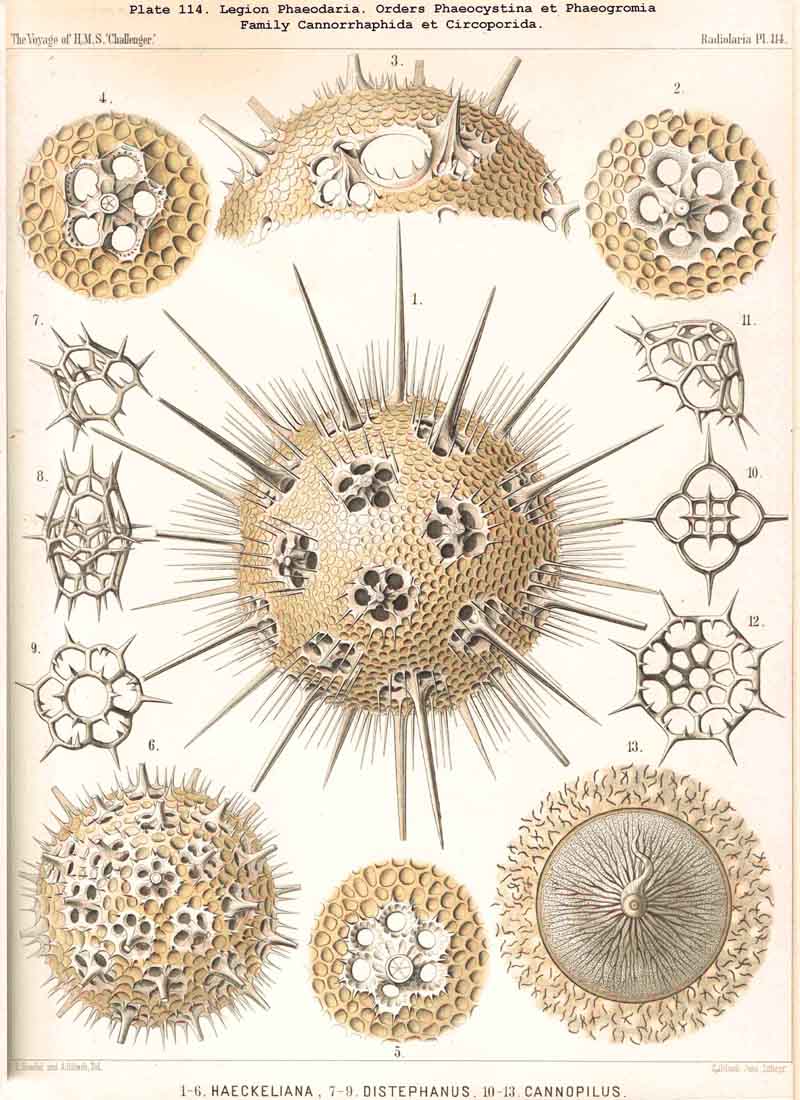
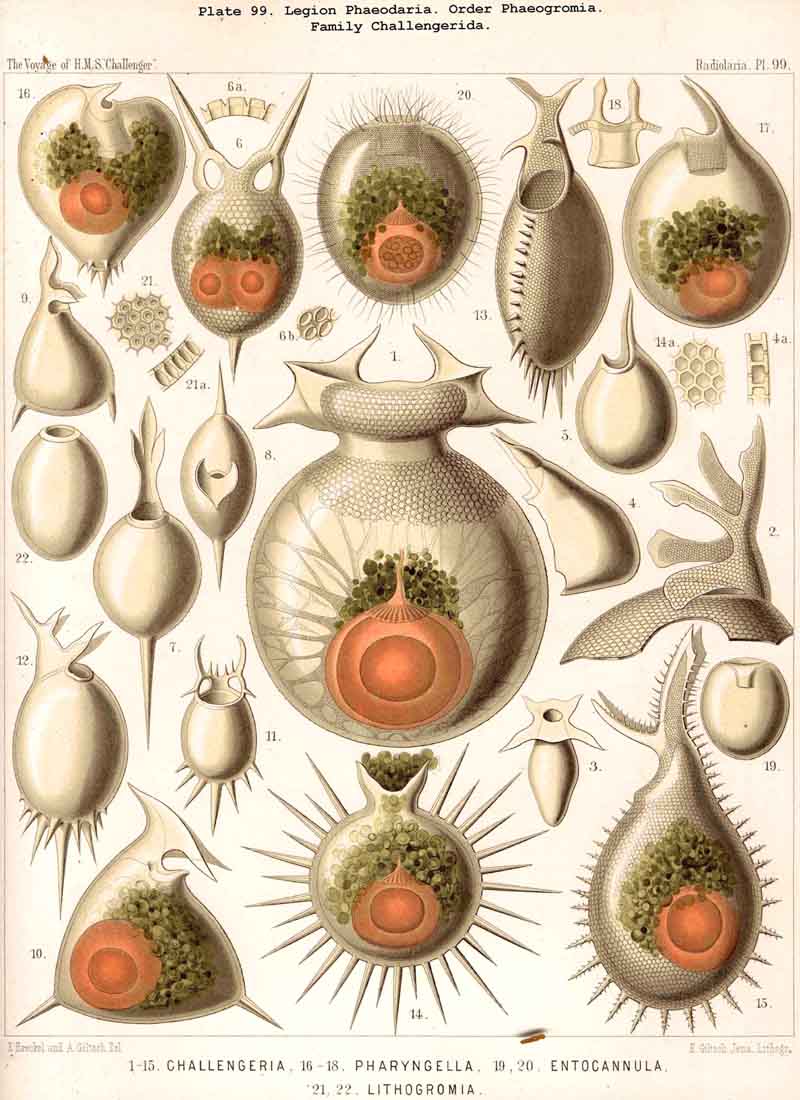
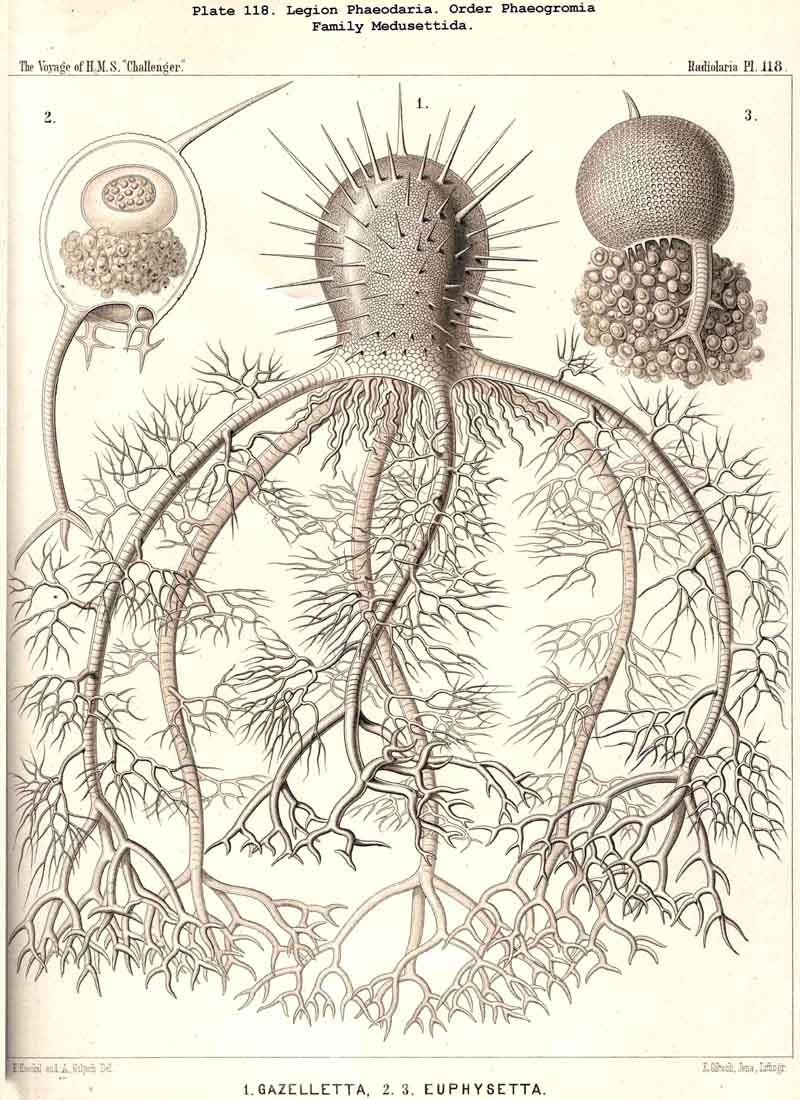
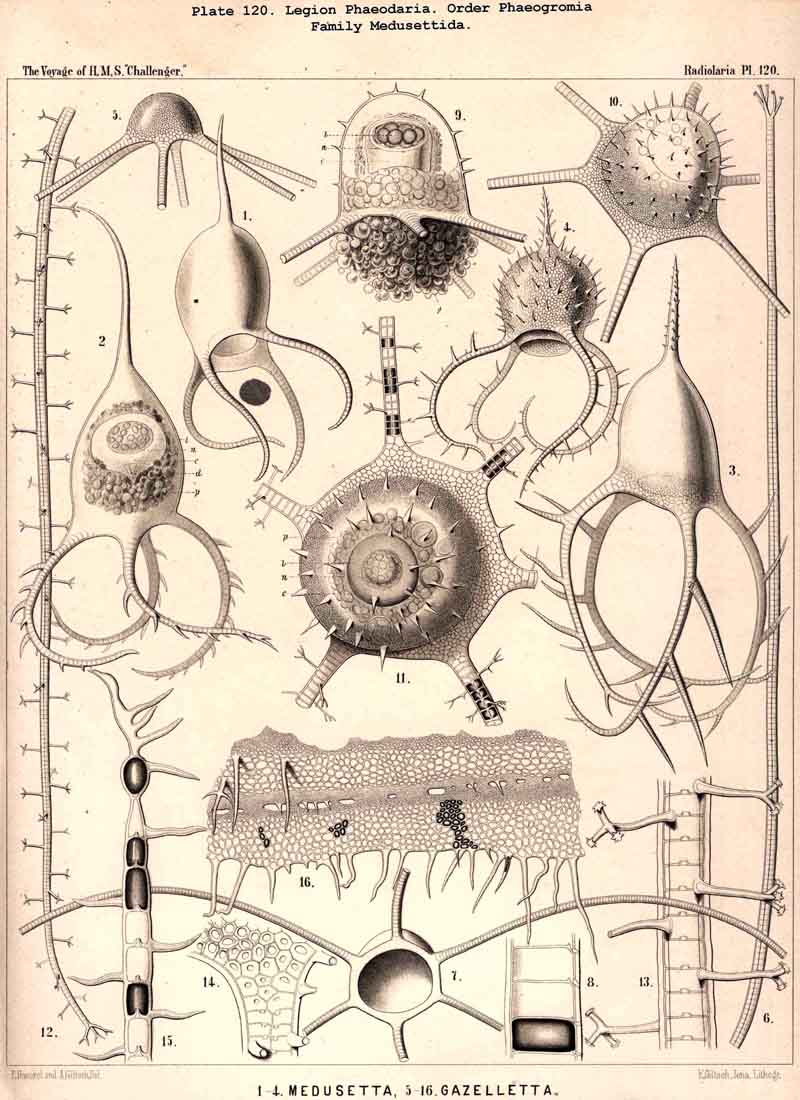
199. Phaeoconchia with a Conchoid Shell.-The order Phaeoconchia (Pls. 121-128) is separated not only from all other PHAEODARIA, but also from all other Radiolaria, by the possession of a bivalved shell resembling that of a Lamellibranch ; the two valves of this Conchoid skeleton are to be interpreted as dorsal and ventral (§ 128). Probably these bivalved shells are independent products, but possibly they may have been formed by the bisection of a simple spherical lattice-shell ; in the former case the Phaeoconchia would be directly descended from the Phaeodinida, in the latter from the Castanellida. The three families which we have distinguished among the Phaeoconchia, probably constitute a connected stem, the most primitive group of which are the Concharida (Pls. 123 ,124, 125). From these the Coelodendrida (Pls. 121, 122). have next arisen by the formation of a "galea" upon the apex of each valve, and the growth of hollow tubes from this helmet-like structure. Finally, the Coelographida (Pls. 120-128) have been developed from the Coelodendrida by the formation of a basal nasal tube (rhinocanna) from each galea, and the formation of a median or paired frenulum, which connects the opening of the nasal tube with the apex of the galea. In the Coelodendricia, as well as in the Coelographida, there are two different subfamilies, of which the more primitive (Coelodorida, Coelotholida) have free branches from the hollow radial tubes, whilst the more recent (Coelodrymida, Coeloplegmida) form an outer bivalved shell by anastomosis of the branches of the tubes.
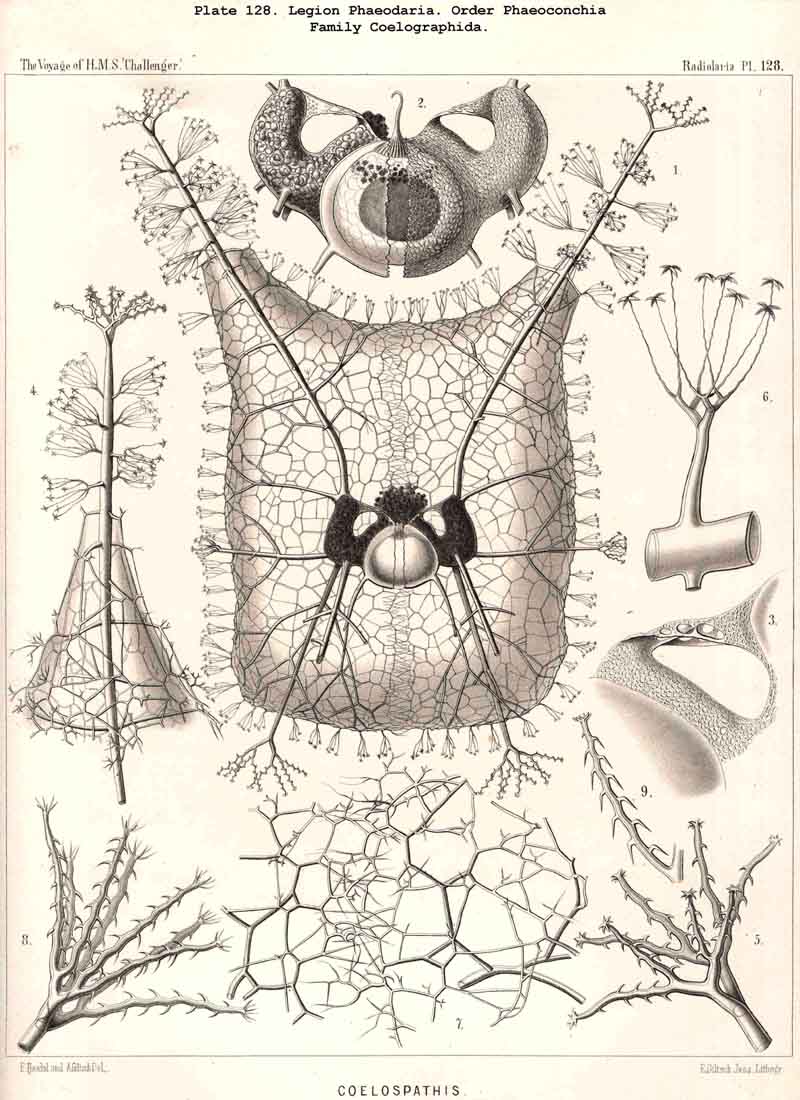
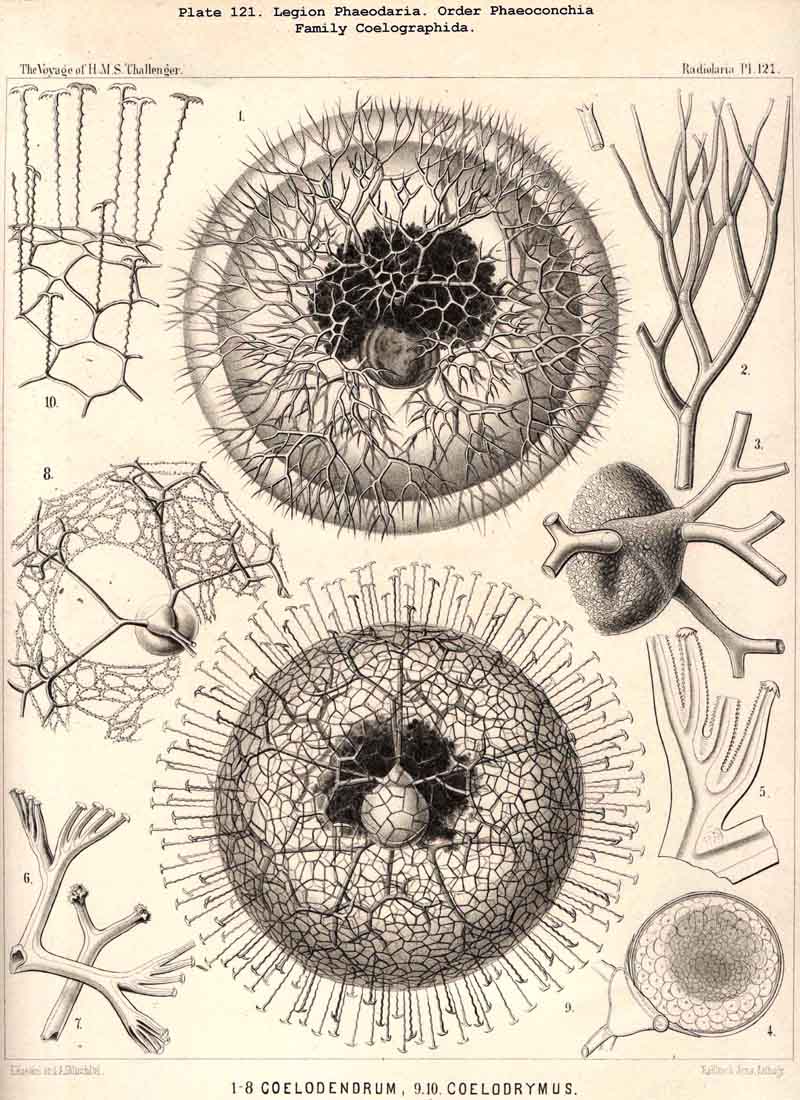
200. The Fundamental Biogenetic Law.-The causal connection between ontogeny and phylogeny, which finds its most precise statement in the fundamental biogenetic law, holds in general for the Radiolaria as for all other organisms. In order to furnish direct proof of this, however, a complete empirical knowledge both of individual and of palaeontobogical development would be necessary. In both these directions, as has been shown in the foregoing chapters, our knowledge of the Radiolaria is very incomplete and fragmentary, but still we are able to convince ourselves indirectly of the validity of the law as applied to Radiolaria by the aid of comparative anatomy. This is now so fully known to us (§§ 1-140) that we are able not only to draw a complete and satisfactory picture of their morphology, but also to arrive at most important conclusions regarding the ontogeny and phylogeny of the individual groups. As regards the formation of the multiform skeleton of the Radiolaria, most of the ontogenetic series of forms, with which we have become acquainted by comparative anatomy, are of palingenetic nature ; that is, they are primarily due to inheritance and thus of direct phylogenetic significance. On the other hand, among the ontogenetic phenomena of the Radiolaria, as far as they have yet been investigated, only very few are cenogenetic, that is, brought about by adaptive modification and without direct significance as regards phylogeny.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
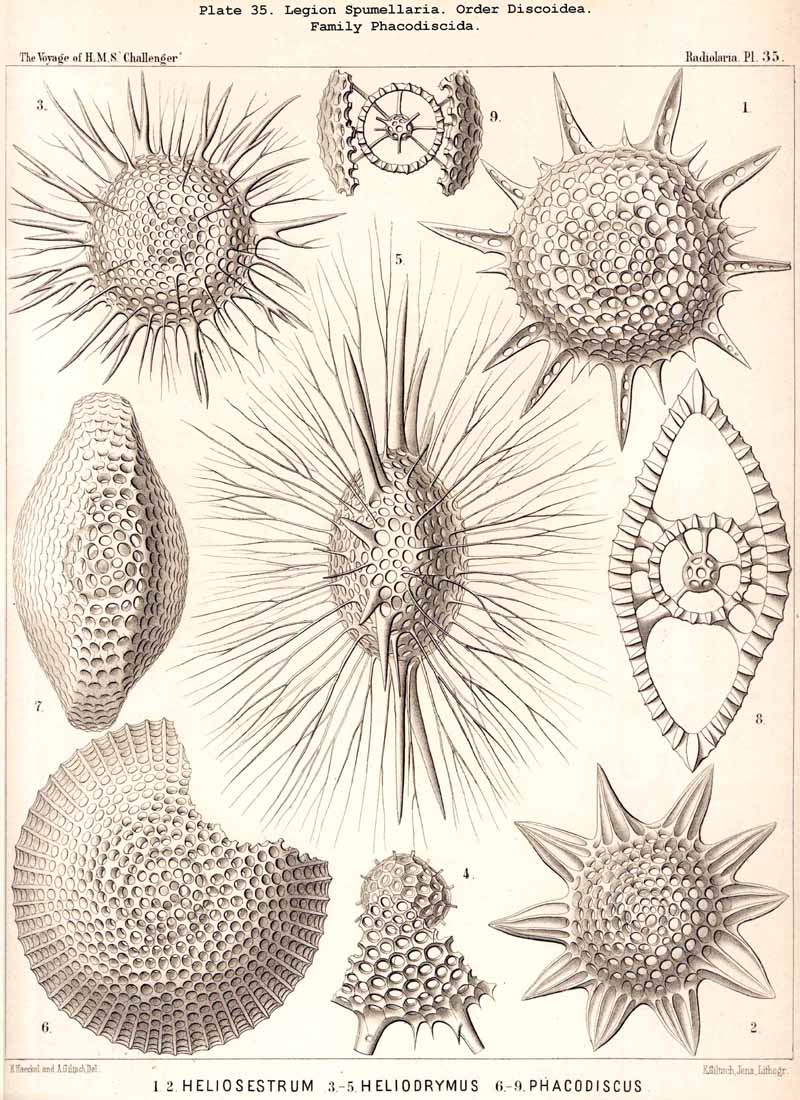
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
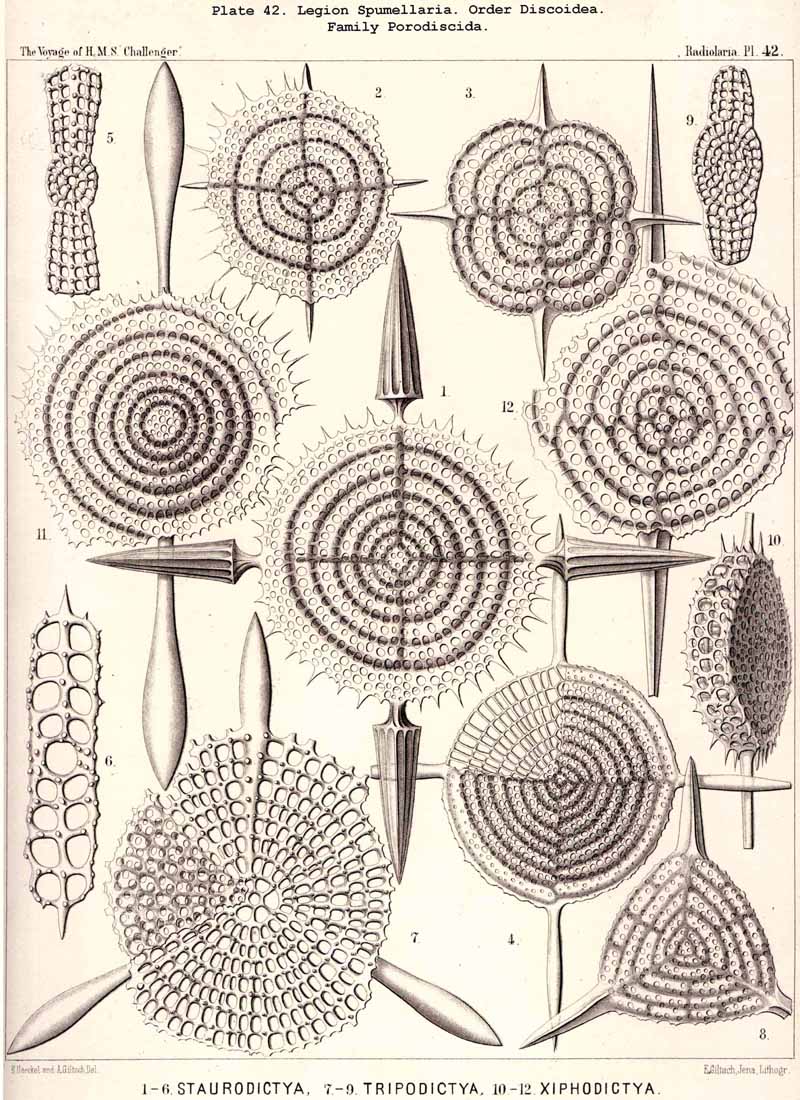
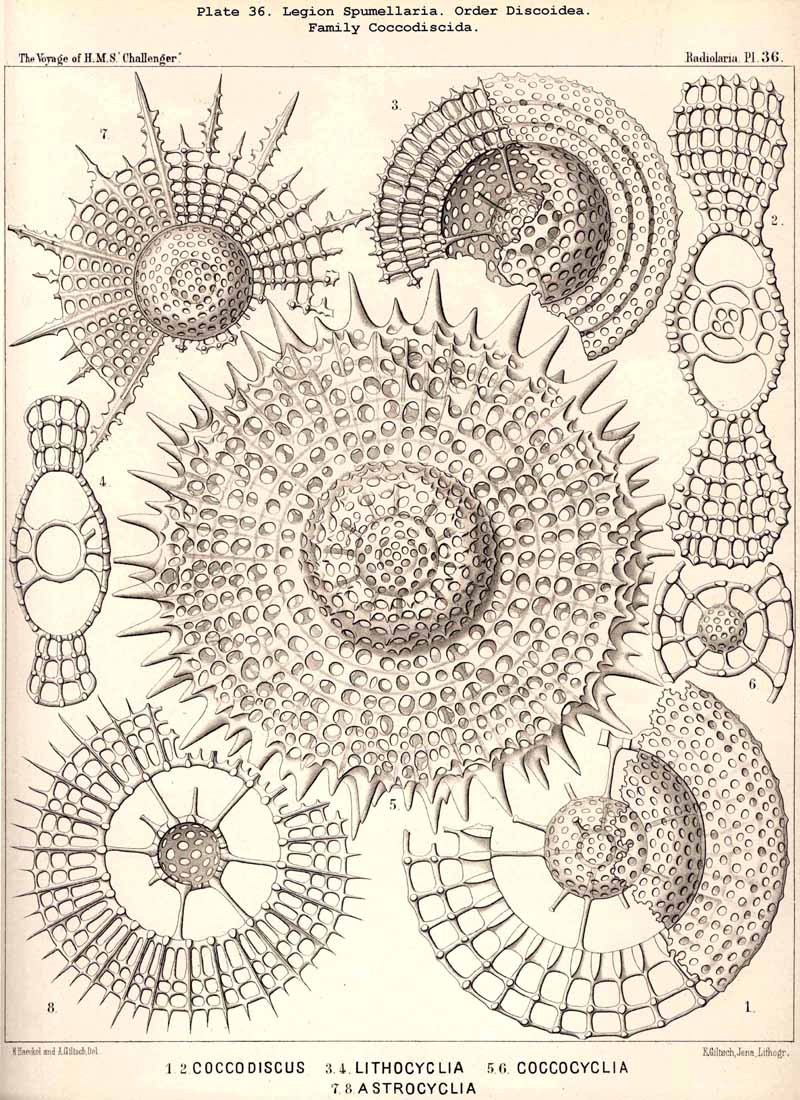
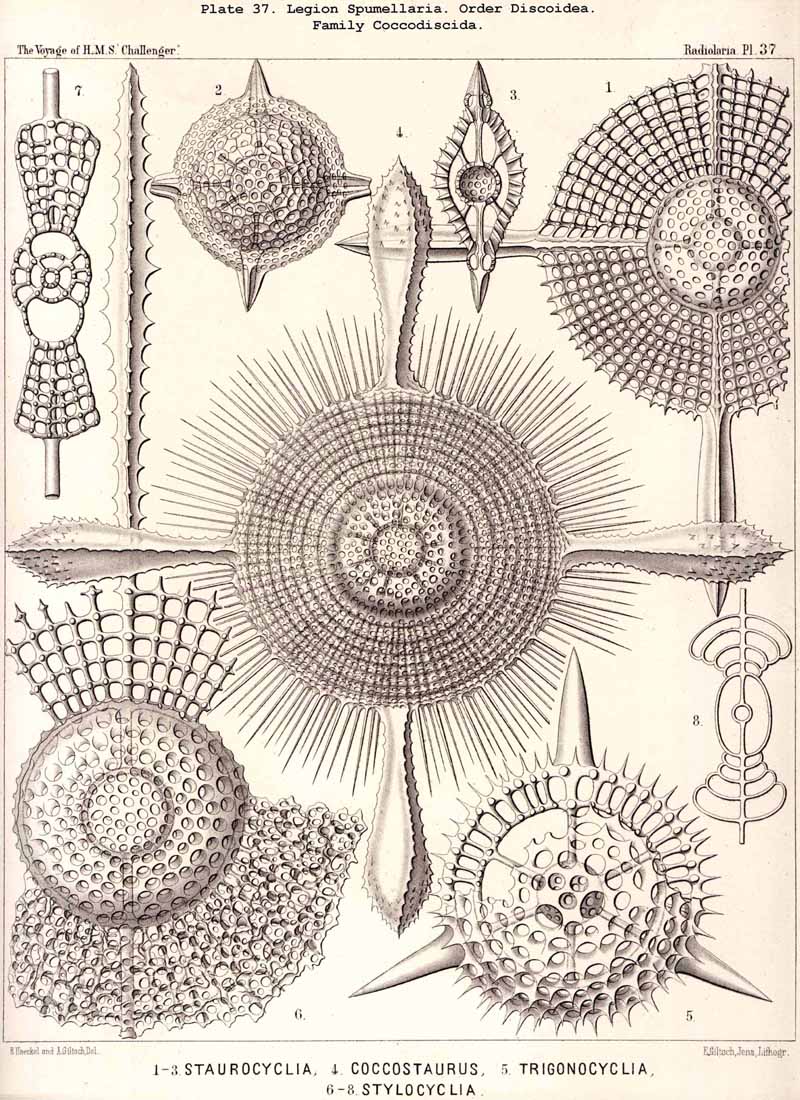
{kind=link}
{kind=link}
{kind=link}
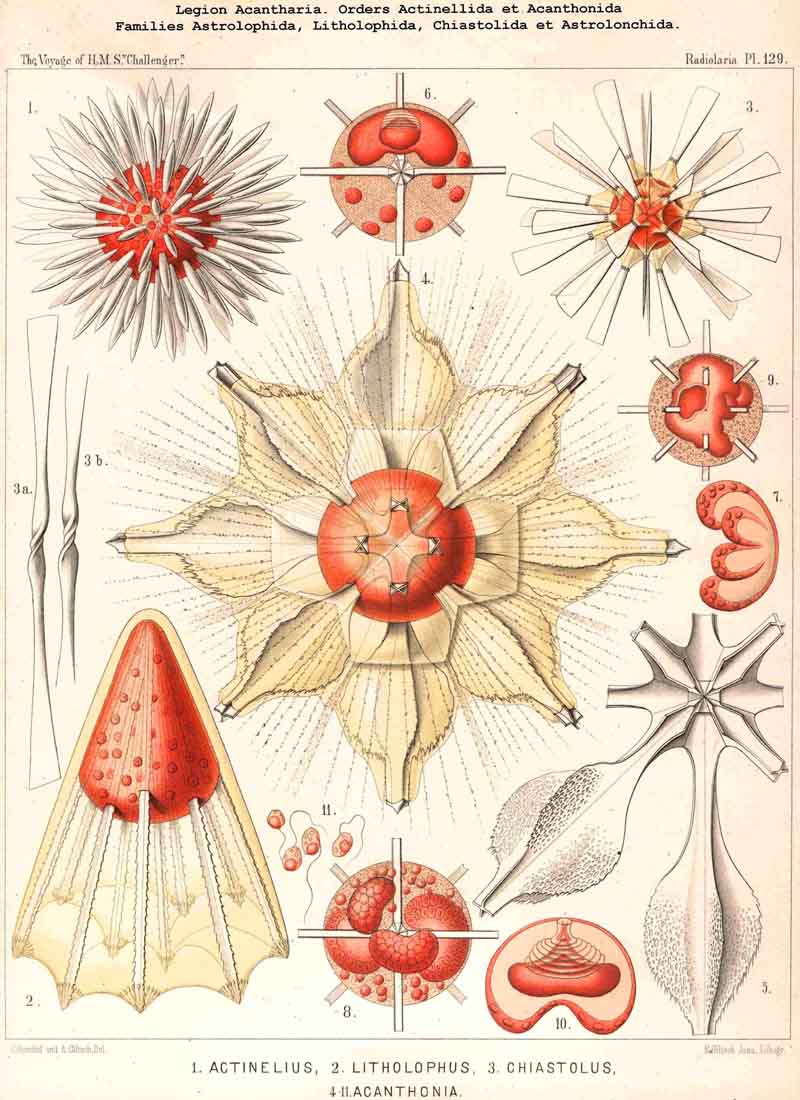
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
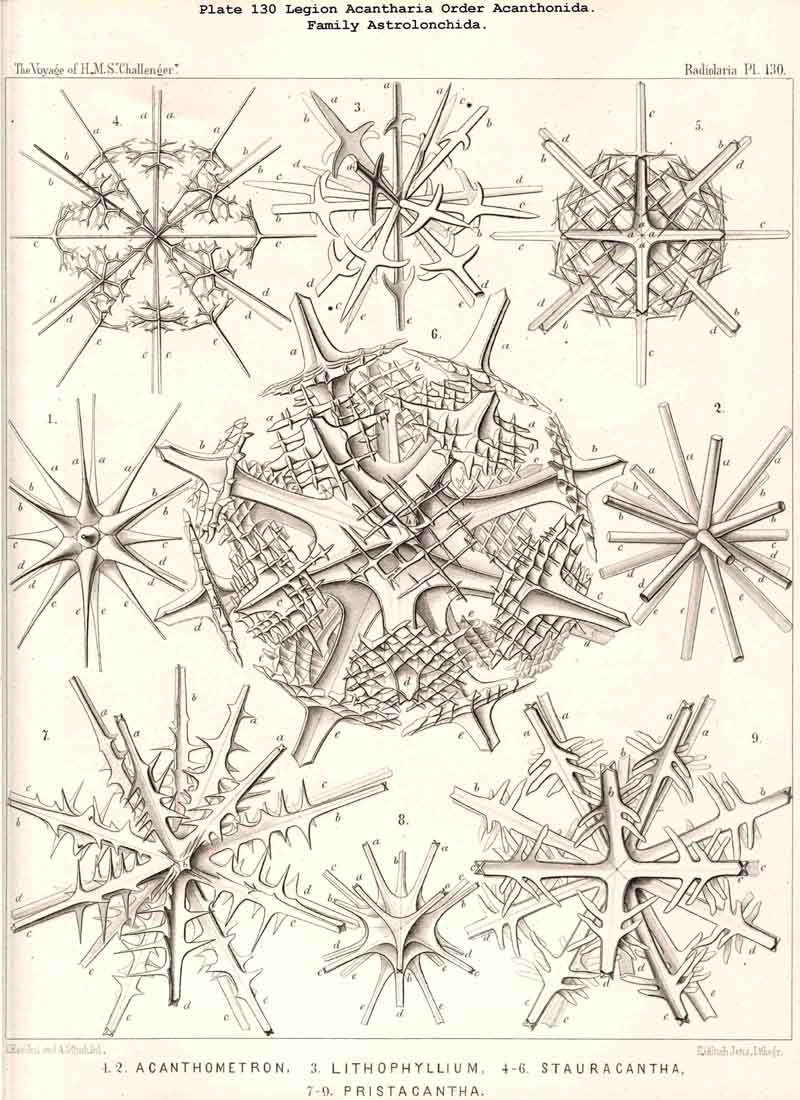
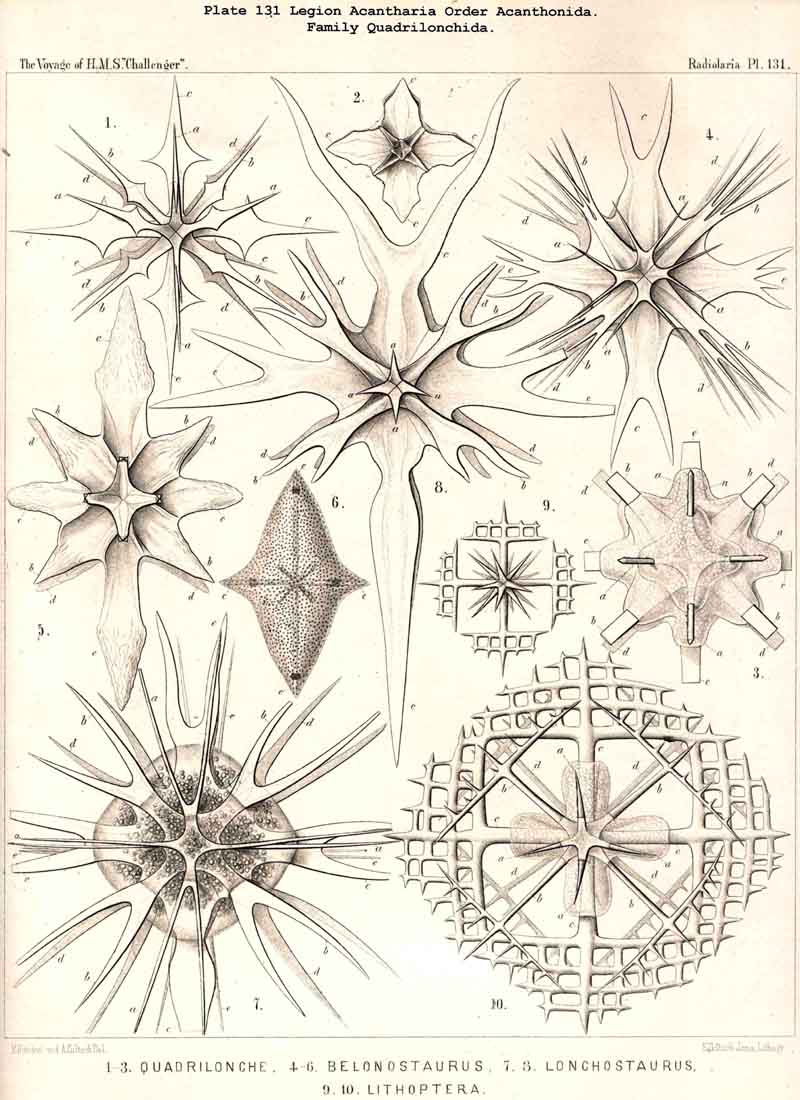
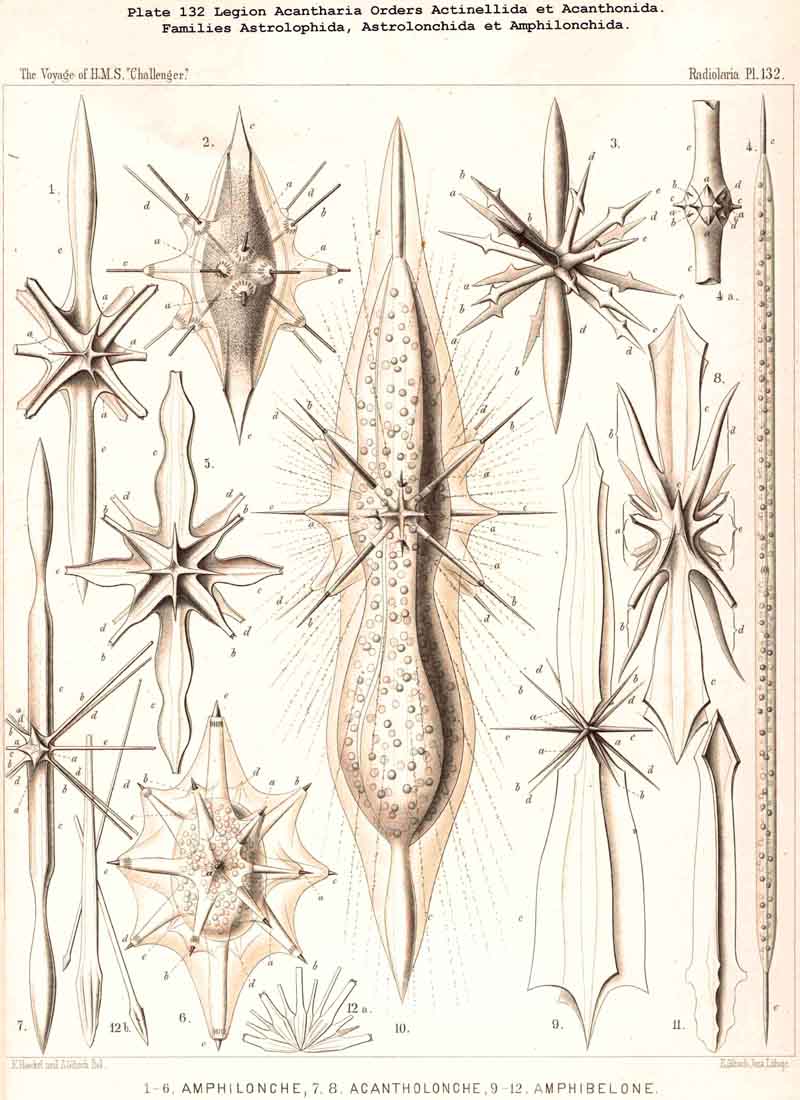
{kind=link}
{kind=link}
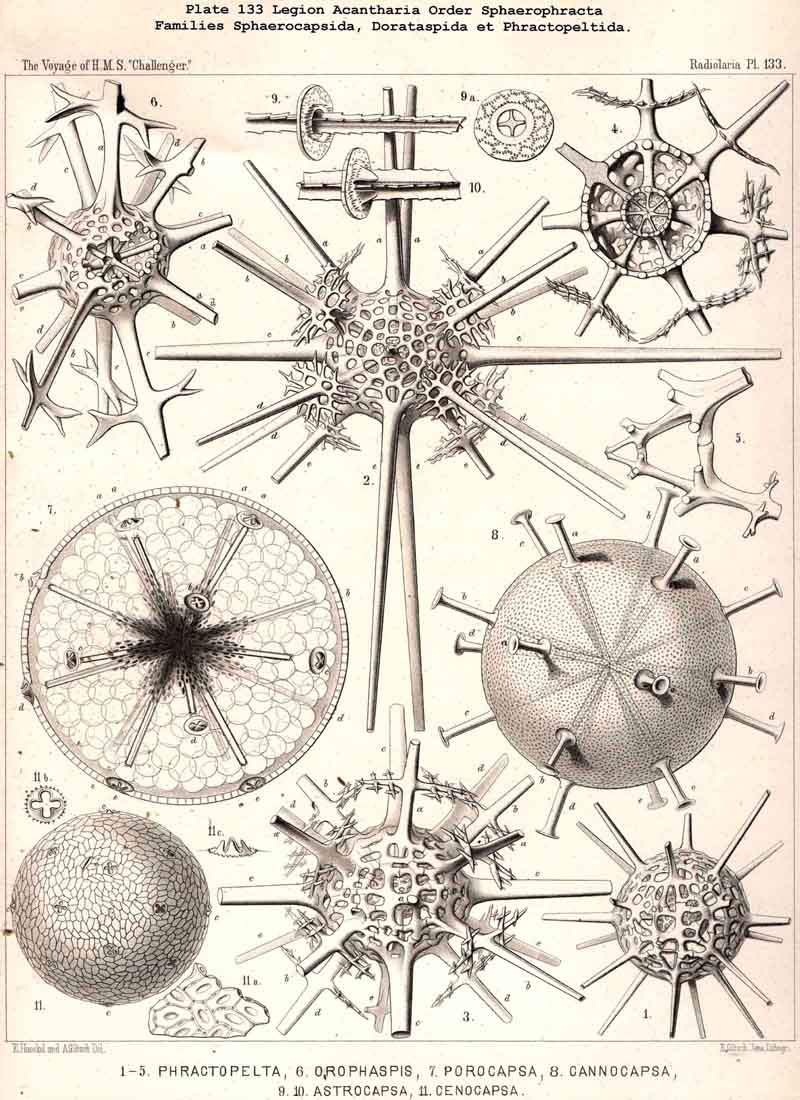
{kind=link}
{kind=link}