81. The Components of the Extracapsulum.-The extracapsulum or extracapsular malacoma, under which name are included all those parts of the soft body which lie outside the central capsule, consists of the following constant and important constituents :-
(1) The calymma or extracapsular jelly-veil; (2) the sarcomatrix or layer of exoplasm immediately surrounding the membrane of the central capsule; (3) the sarcodictyum or network of exoplasm, covering the surface of the calymma; (4) the pseudopodia or radial fibres of exoplasm, which may again be subdivided into intracalymmar pseudopodia, uniting the sarcomatrix and sarcodictyum, and extracalymmar pseudopodia, radiating freely into the water outside the calymma.
82. The Calymma.-The calymma or extracapsular jelly-veil of the Radiolaria is always the most voluminous portion of the extracapsulum, and in spite of its simple structureless constitution is of great morphological and physiological importance. In all Radiolaria this gelatinous mantle completely surrounds the central capsule, but is separated from its outer surface by a continuous, though thin, layer of exoplasm, the sarcomatrix. The pseudopodia radiating from the latter pierce the calymma, form the sarcodictyum at its surface, and radiate from its nodal points freely into the surrounding water. The calymma is rarely visible in living freshly captured Radiolaria, examined in sea-water, for its gelatinous substance is perfectly hyaline, colourless and pellucid, and possesses the same refractive index as sea-water; but when the object is removed from this fluid and transferred to carmine solution or some other colouring matter, the extent and figure of the calymma become apparent, for the staining fluid does not at first penetrate into the gelatinous material. When this has taken place, however (after a longer or shorter time), and the gelatinous material has become coloured, its form and size may be observed by the converse experiment; the object is transferred once more to water and the outlines of the calymma become as clear as those of the central capsule. The same is the case with dead specimens in which the sticky surface of the calymma has become covered with dust.
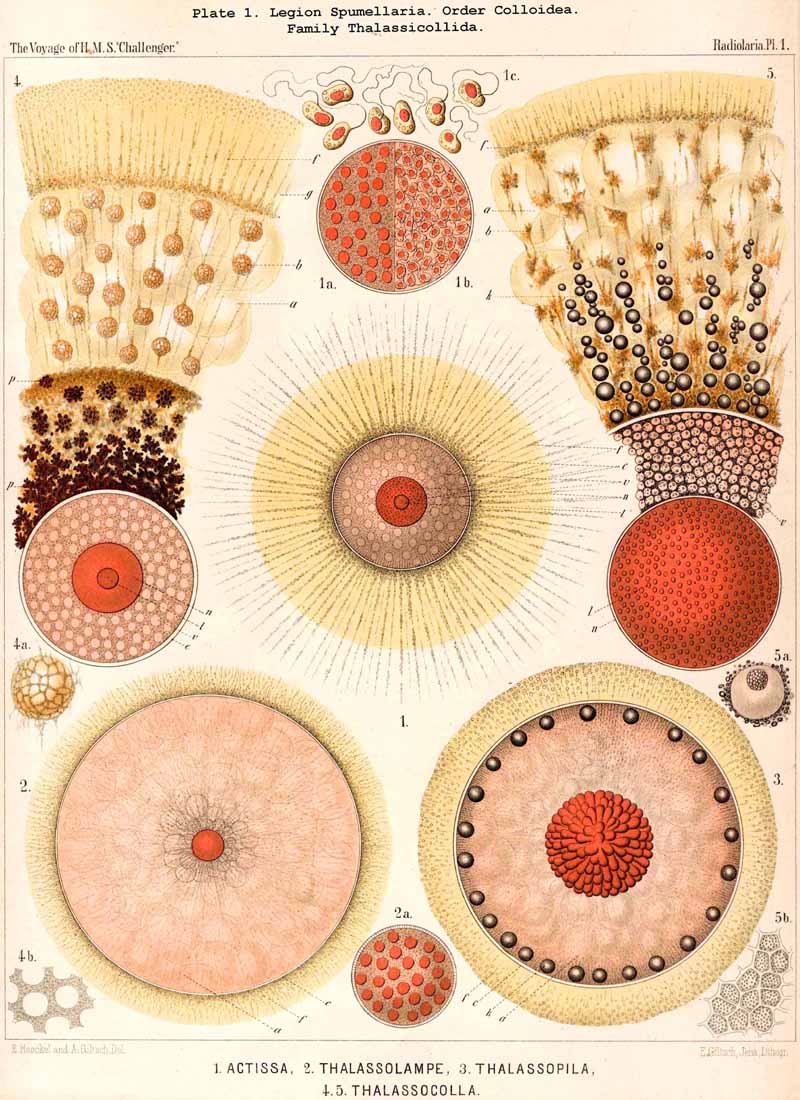
The jelly-veil of the Radiolaria was recognised even by the earliest observers of the group, Meyen (1834), and Huxley (1851), and compared with that of the Palmellaria; the former noticed it in Physematium and Sphaerozoum (L. N. 1, p. 283), and the latter in Thalassicolla and Colbosphaera (L. N. 5, p. 433). In all these SPUMELLARIA, both in the monozootic Thalalassicolla and in the polyzootic Sphaerozoum and Collosphaera, the calymma is very voluminous and filled with large alveoli. Meyen called them "muco-gelatinous masses, in the interior of which are contained small equal-sized vesicles"; Huxley likewise found clear vesicles in the jelly and compared them with Dujardin's vacuoles. Johannes Müller observed the jelly-veil in many different Radiolaria, in particular in the Acanthometra, first discovered by him, but erroneously believed that it only originated after death by liquefaction of the sarcode (L. N. 12, p. 6). This mistake is, however, easy to understand, since in living Radiolaria the calymma is usually invisible on account of its perfect transparency, whilst in dead specimens it is usually quite distinct on account of the dust clinging to its adhesive surface. I myself believed that the formation of the voluminous hyaline jelly-veil was only partially due to liquefaction after death, but that it was to some extent present in the living organism and that it might vanish and subsequently reappear by means of imbibition (L. N. 16, pp. 109, 110). R. Hertwig was the first to demonstrate, in 1879, that the jelly-veil is constantly present in living Radiolaria, that it forms the basis of the extracapsular malacoma and surrounds the central capsule as a second protective sheath (L. N. 33, p. 114).
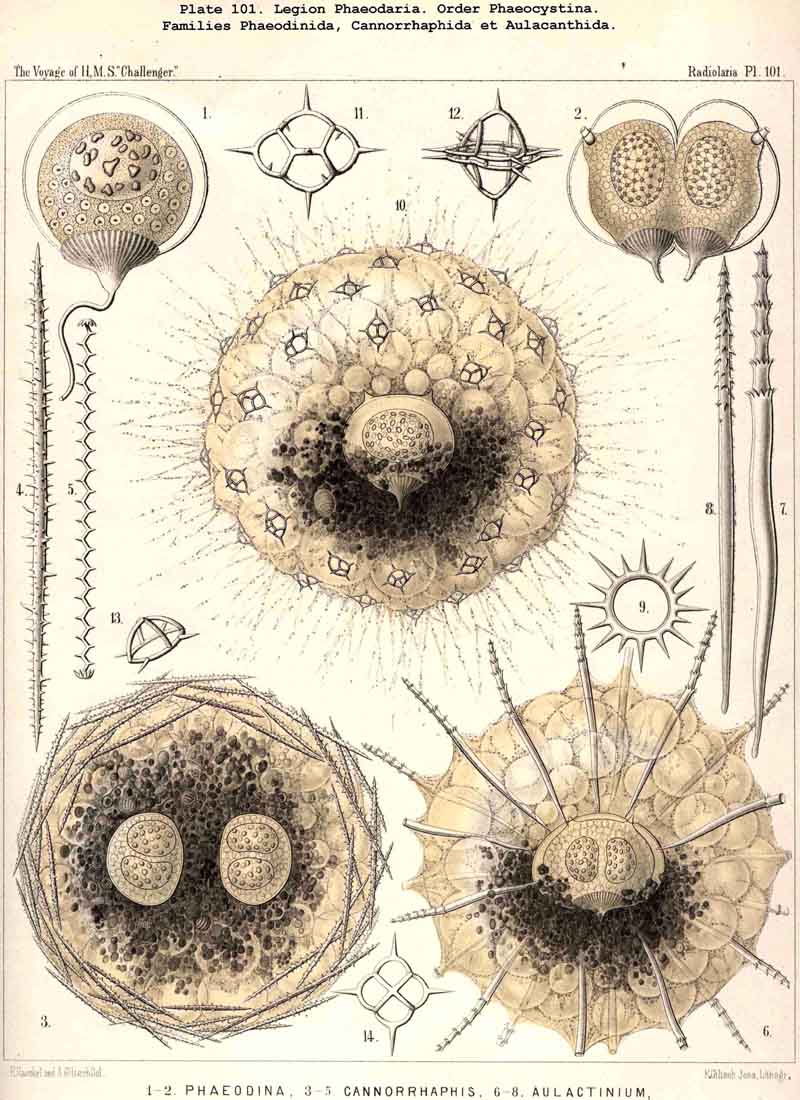
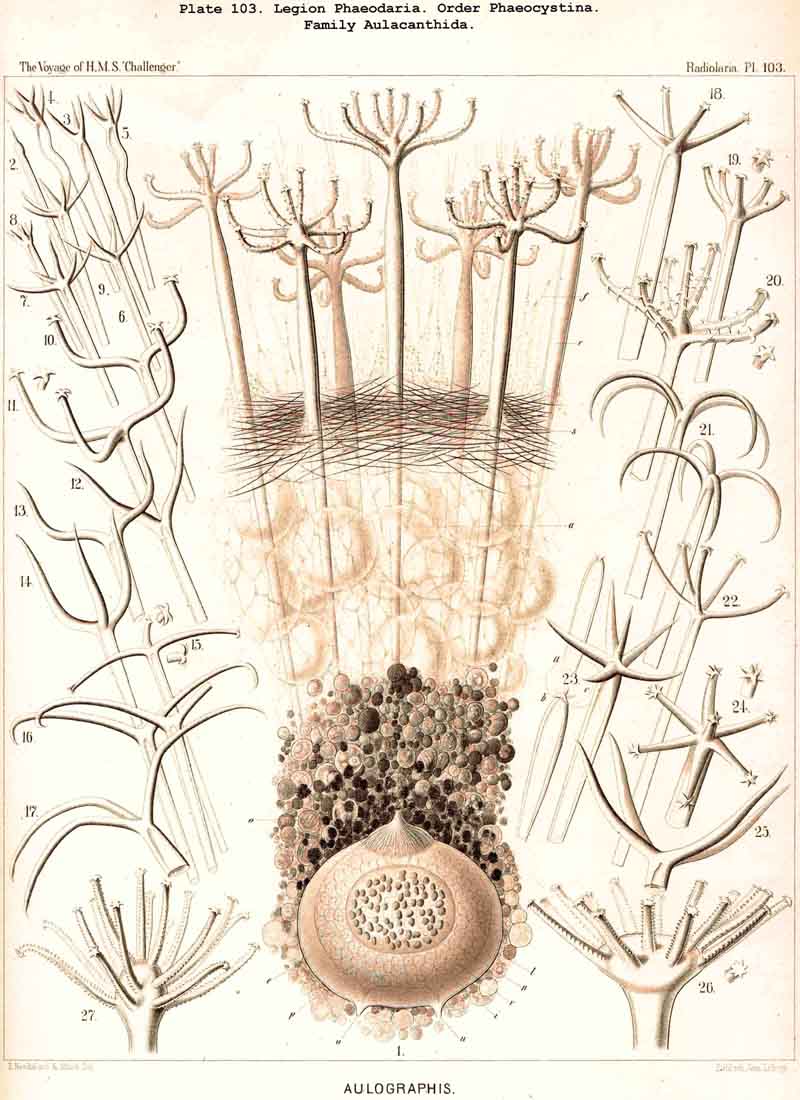
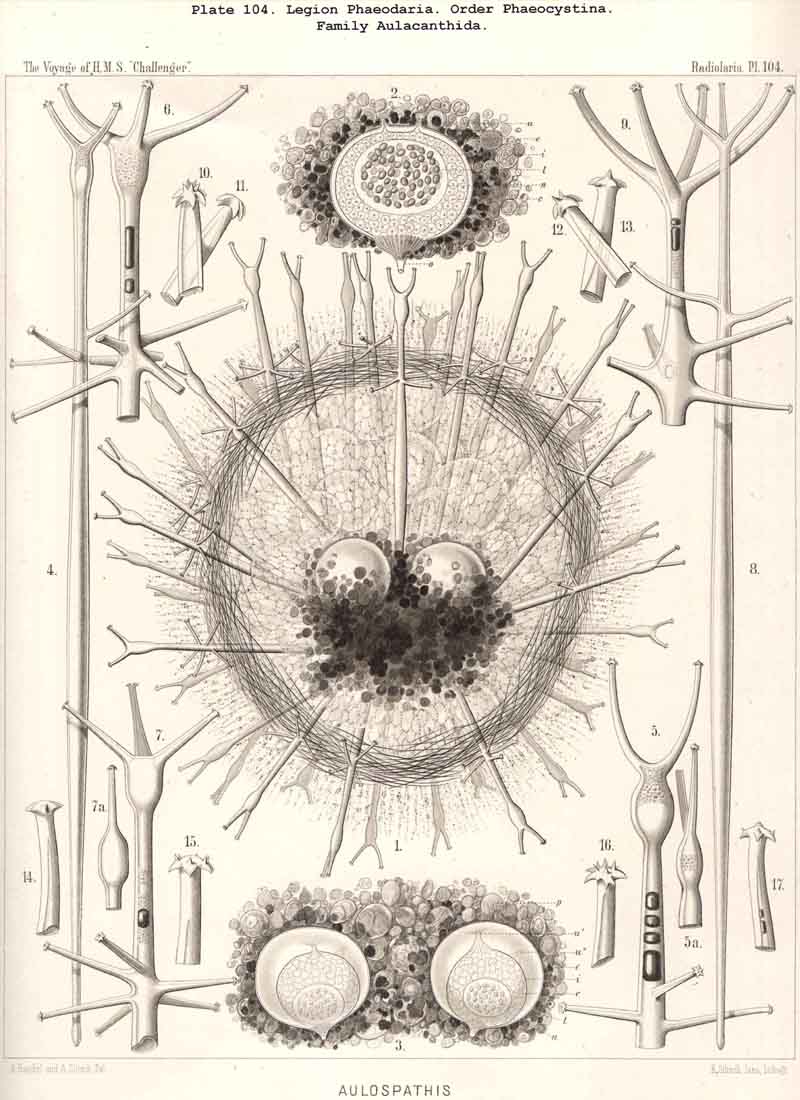
83. The Structure of the Calymma.-The extracapsular jelly-veil appears structureless in most Radiolaria, inasmuch as it represents a homogeneous pellucid excretion of the exoplasm and contains neither fibres nor other formed structures. In some groups, however, definite structural characters become secondarily developed. The most common and striking of these is the formation of alveoles, which takes place in the extracapsulum (see § 86). In consequence of this the calymma assumes a remarkable frothy consistency and appears to be composed of large, clear, thin-walled vesicles; this is especially the case in the Collodaria (Colloidea, Pls. 1,3, and Beloidea, Pls. 2,4), and in many large PHAEODARIA, especially among the Phaeocystina (Phaeodinida and Cannorrhaphida, Pl. 101, and Aulacanthida, Pls. 102, 103,104). More rarely the calymma is not permeated by vacuoles, but there appear in it fine striae parallel to the surface as though it were composed of thin concentric laminae like an onion; perhaps these are the expressions of a different quantity of water in the various layers. In the calymma of many Radiolaria thin, straight, radial lines are to be seen, which are probably pseudopodia, and not to be attributed to any structural modification, or they may be slender canals which serve for the exit of the pseudopodia. On the outer surface of the calymma of different Radiolaria, and especially in the ACANTHARIA, a peculiar network of fibres is to be found, composed of polygonal meshes, like elastic fibres, probably due to a local thickening of the jelly. These polygonal meshes are often very regularly distributed between the radial spines of the Acanthometra, and stand in a definite relation to them. The fibres which form the meshes are often rather strong, resembling elastic fibres, as above-mentioned, and either simple or composed of bundles of very fine fibrillae (L. N. 33, p. 15, Taf. i. fig. 1, Taf. ii. fig. 4).
84. The Consistency of the Calymma.-The gelatinous material of which the calymma of the Radiolaria consists is a pellucid mass, rich in water and usually quite hyaline and structureless; its consistency is very variable. In the majority of the Radiolaria it may perhaps be about equal to that of the jelly which composes the umbrella of most Medusae; but as in these latter it may vary between very wide extremes, constituting on the one hand a very soft jelly-mantle, offering but little resistance to mechanical influences and almost disintegrating under the eyes of the observer, and on the other hand forming a firm gelatinous shell, comparable to cartilage in hardness, elasticity, and power of mechanical resistance. In many Radiolaria of large dimensions with an alveolar calymma (especially in numerous Collodaria and PHAEODARIA) this may be split by means of dissecting needles and the central capsule extracted like the stone from a cherry, and then it is easy to ascertain that the firmness and elasticity of this jelly-veil are not less than those of a cherry. The different degrees of consistency in the various Radiolaria may be dependent either upon the relative amount of water which they contain, or upon qualitative or quantitative variations in the organic substance of which the jelly consists. Great importance is to be attached to the considerable consistency of the calymma, because it furnishes the indispensable groundwork for the deposition of many parts of the skeleton and particularly of the lattice-shells.
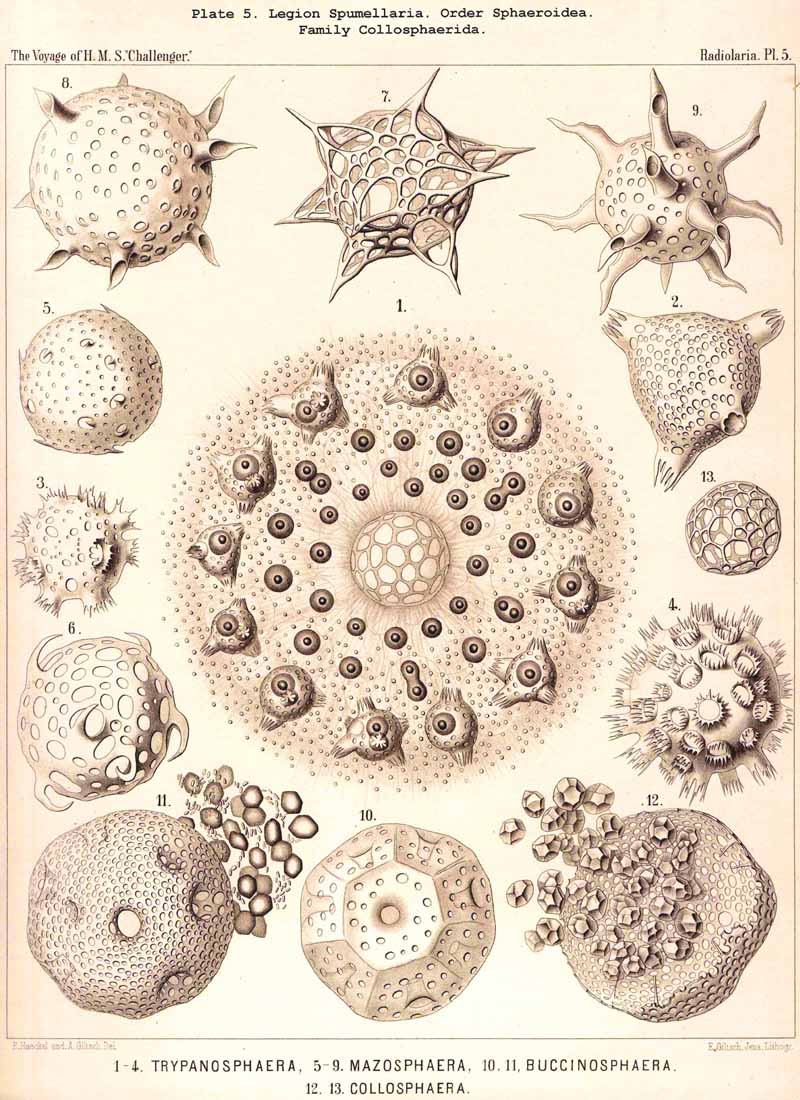
85. The Primary and Secondary Calymma.-In most Radiolaria the external form and volume of the calymma are different at different stages of growth, and this difference is mainly dependent upon the development of the skeleton. Hence it is ad visable to distinguish in general the primary from the secondary calymma. The primary calymma is in the great majority of Radiolaria a perfect sphere, in the middle of which lies the concentric central capsule; on the surface of this gelatinous plate the primary spherical lattice-shell is secreted in most SPUMELLARIA and Acanthophracta, as well as in those PHAEODARIA which possess a spherical shell; in the remaining PHAEODARIA also and in the NASSELLARIA, where the lattice-shell is not spherical but monaxon, it is secreted on the surface of the primary calymma. This takes place at a definite time, very important in the development of the Radiolarian, which for the sake of brevity we shall term the "lorication-period." Since the firm surface of the primary calymma furnishes the necessary foundation for the deposition of the primary lattice-shell, it is of the greatest mechanical significance in all shell-bearing Radiolaria. The secondary calymma arises only after the lorication-period by further growth of the primitive jelly-mantle and in the fully developed Radiolarian usually encloses wholly or partially the external parts of the skeleton, in consequence of which it assumes the most various forms. Very often the secondary calymma is polyhedral, being stretched between the radial spines of the skeleton, the distal ends of the latter then forming the fixed points of the gelatinous polyhedron.
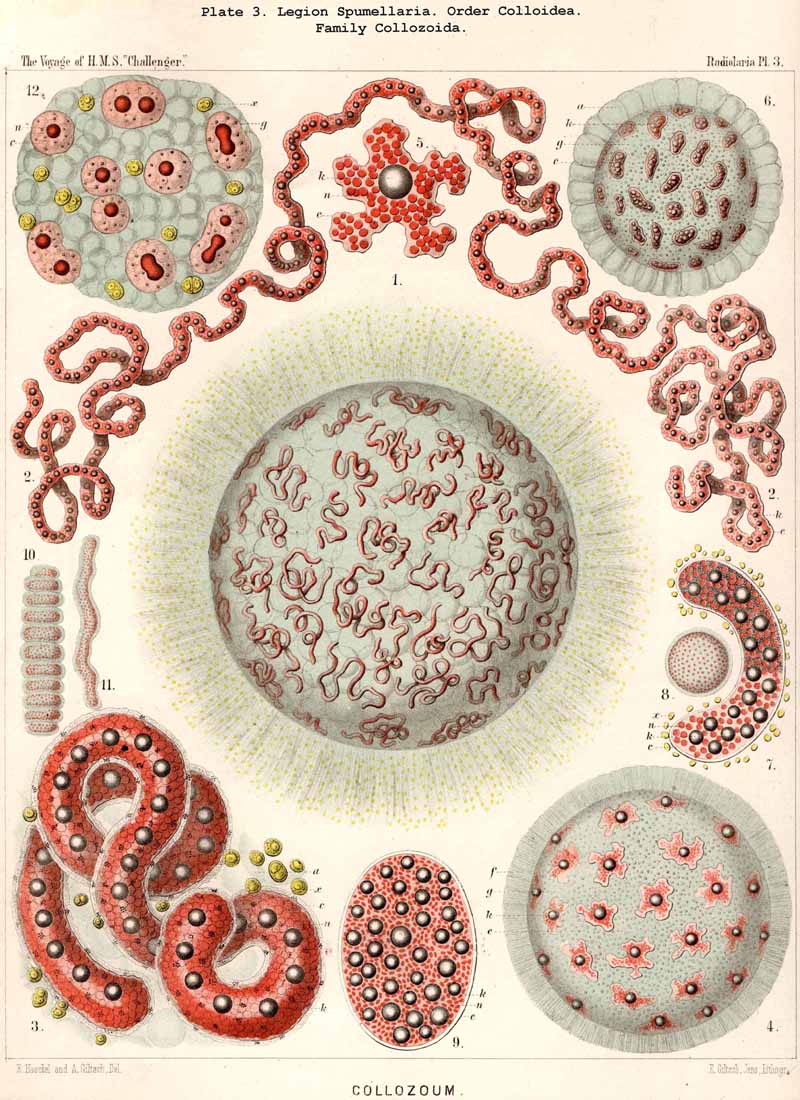
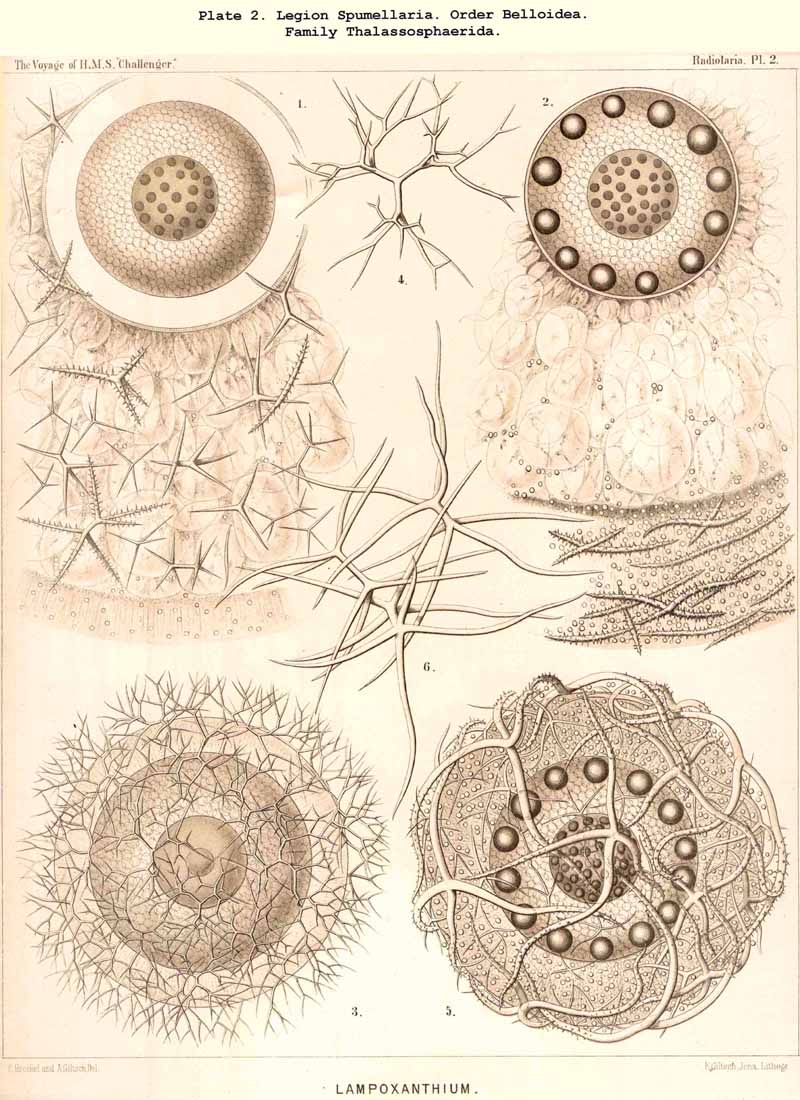
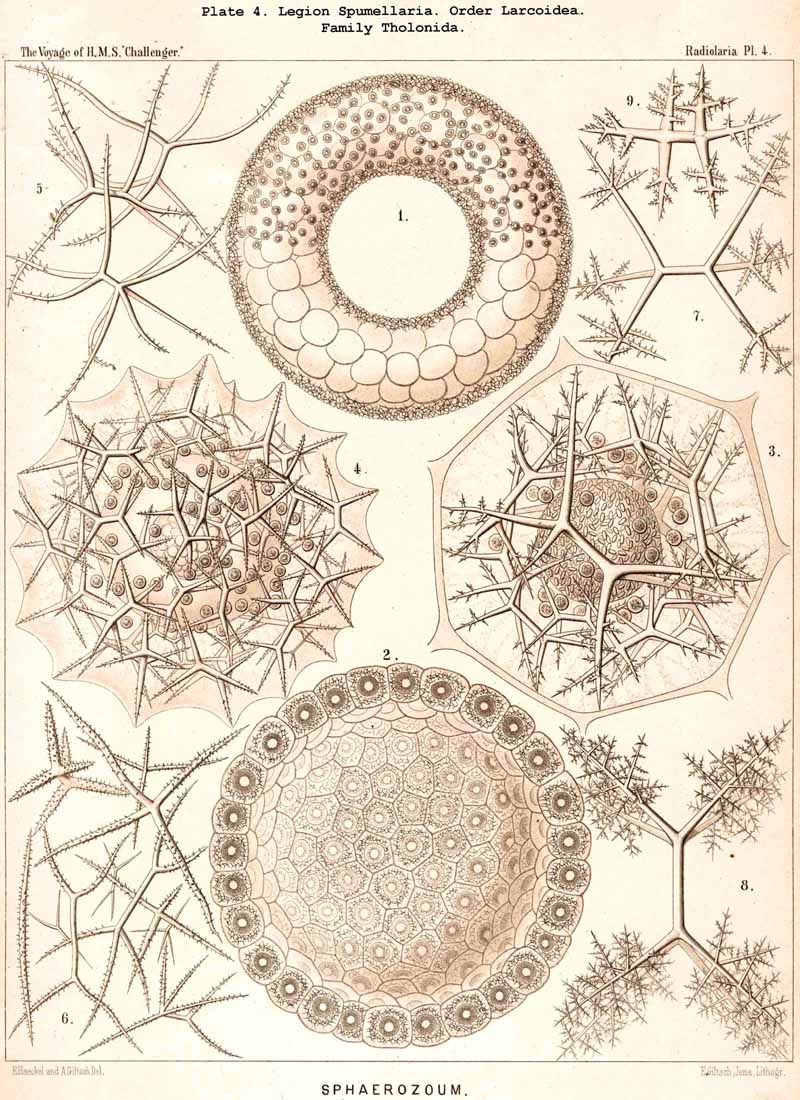
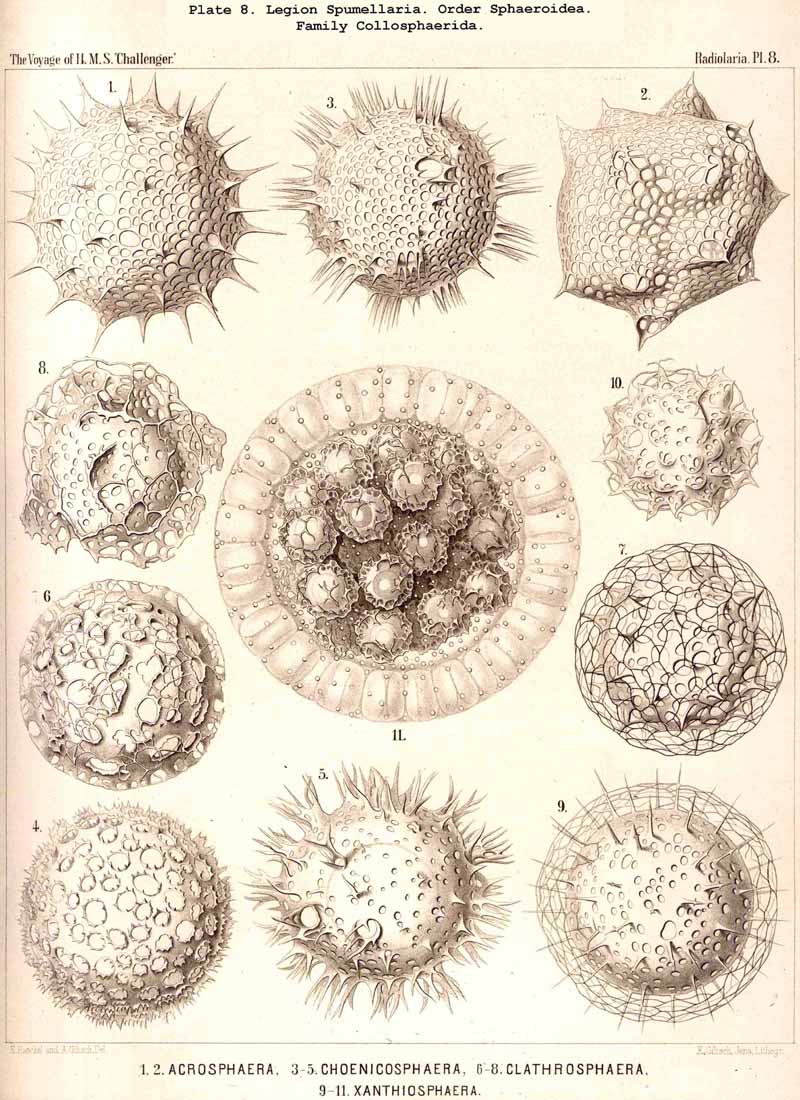
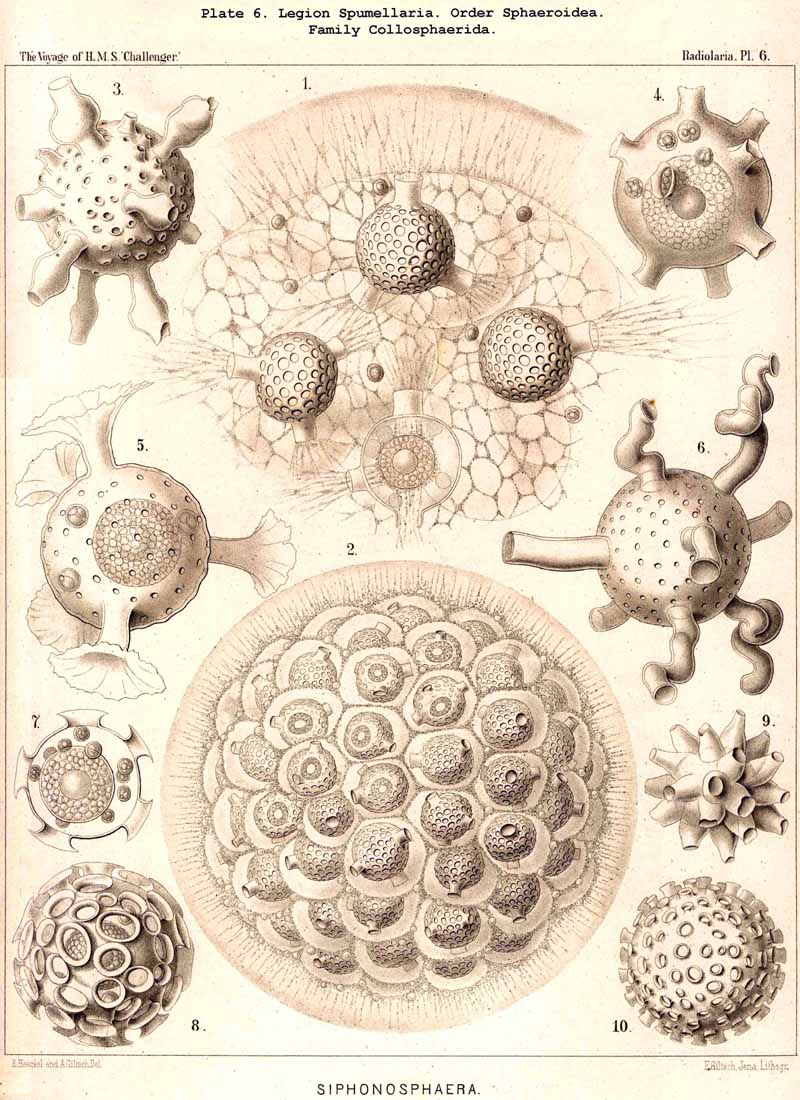
86. The Extracapsular Vacuoles and Alveoles.-The calymma of the Radiolaria usually appears completely homogeneous and hyaline without any structure; some times it encloses numerous clear vesicles, vacuoles or alveobes, and then assumes a frothy appearance, the expression of a more or less distinct alveolar structure. The clear vesicles to which this is due are either spherical, or polyhedral from mutual pressure, and like the similar ones in the central capsule may be divided into membrane-less vacuoles and vesicular alveoles. The vacuoles are simple drops of fluid, without a special envelope, and immediately surrounded by the gelatinous substance of the calymma, in which they appear as simple cavities. The alveoles on the contrary are true vesicles, with a thin envelope, which encloses a drop of fluid or a globule of jelly; in the latter case its contents are different in refracting power and amount of contained water from the substance of the surrounding calymma. A sharp boundary between the membraneless vacuoles and the vesicular alveoles cannot be drawn in the case of the extracapsular hyaline spheres any more than in the intracapsular; the envelope of the alveoles is sometimes very distinct and even anatomically separable, whilst at other times it is very thin and scarcely recognisable; it may occasionally arise and disappear within a very short time (see note A). There is no doubt that in the calymma as in the central capsule the vesicular alveoles are secondary products, which have arisen from the vacuoles by the secretion of an enveloping membrane. This membrane is either a delicate sheath of exoplasm, or a firmer and more resistant skin, distinct from the exoplasm, and probably an excretion from it (e.g., Pl. 4, figs. 2, 3). In many cases the outer surface even of the vacuoles is covered by a network of pseudopodia, which form a sarcoplegma similar to a fenestrated alveolar membrane. The colourless pellucid fluid in the vacuoles and alveoles is usually simple sea-water, more rarely it contains a small quantity of albumen ("albumen-spheres") or jelly ("gelatinous spheres"). The size of these spheres is very variable. Quite small vacuoles may be found in the calymma of many Radiolaria. Large vacuoles, on the other hand, producing the appearance of an alveolar structure, are confined to but few groups, to a part of the SPUMELLARIA (Colloidea, Beloidea, and a few Sphaeroidea), and to the Phaeocystina (PHAEODARIA with incomplete skeleton); besides they occur only rarely in individual genera, e.g., Nassella among the skeletonless NASSELLARIA. Since the volume of the calymma is much increased by the development of vacuoles, and the power of mechanical resistance is at the same time much increased, the fact is explained that the vacuoles occur mainly in Radiolaria which have no skeleton or only an incomplete one (see note B). Among the monozootic Colladaria the alveolar structure is especially well developed in the following genera; Thalassicolla (Pl. 1, figs. 4, 5), Thalassophysa, Thalassoplancta, Lampoxanthium (Pl. 2, figs. 1, 2); among the PHAEODARIA in most genera of the Phaeodinida, Cannorrhaphida and Aulacanthida (Pls. 101, 102, 103, 104), and probably also in other voluminous PHAEODARIA (e.g., Phaeosphaeria). The alveoles or vacuoles in the calymma of these large Radiolaria lie usually in several layers, one above another, and increase in size from within outwards. The Polycyttaria or social Radiolaria (the three families Collozoida, Sphaerozoida and Collosphaerida) without exception have an alveolar structure, and the special form of their colonies or coenobia is to a great extent determined by the development, number, size and arrangement of the alveoles in their calymma (compare Pls. 3-8). In these cases there is not unfrequently developed a large central alveole (see note C) whose thickened wall encloses a globe of jelly and serves as the central support of the whole colony (Pl. 5, fig. 1). Still more striking, however, is the arrangement in certain Polycyttaria, where each individual of the colony (or each central capsule with its calymma) is enclosed in a large alveole, whose firm wall often attains considerable thickness (Pl. 4, figs. 2, 3). The whole colony then appears as an aggregate of numerous cells, each of which possesses two envelopes, the inner central capsule and the outer alveolar membrane; between these lies in the Collosphaerida the siliceous lattice-shell (Pl. 6, fig. 2). These pericapsular alveoles may be regarded as an outer cell-wall more correctly than the membrane of the central capsule itself, but the arrangement may also be compared to the temporary encystation of other Protista (see note D).
A. The extracapsular vacuoles in the calymma were first observed in 1851 by Huxley, in Thalassicolla and Sphaerozoum, and compared with Dujardin's sarcode vacuoles (L. N. 5). After wards J. Müller noticed that generally these "large clear vesicles are covered by a fine membrane," and hence he called them "alveoles" (L. N. 12, pp. 3, 7, &c.). In my Monograph I have described them more in detail as "extracapsular alveoles" (1862, p. 88, Tafs. i.-iii. xxxii.-xxxv.). Ever since then the point has been debated whether these clear spaces are simple vacuoles in the sense of Huxley or vesicular alveoles as stated by J. Müller. This contention is unnecessary, for both varieties are present, and often no sharp line can be drawn between them. R. Hertwig has recently come to the conclusion that they are as a rule "membraneless vacuoles," but that they "sometimes become surrounded by a special envelope" (L. N. 33, p. 31). He even succeeded "in extracting from a Collosphaera the large vesicle which lies in the centre of many colonies and removing its covering of central capsules and jelly."
B. The mechanical importance of the alveolar structure, which certainly increases the elasticity and mechanical resistance of the voluminous calymma, has not yet been sufficiently realised; in the case of those Radiolaria which have no skeleton, or at all events no lattice-shell, it may take the place of this as a protective envelope. Furthermore, by taking in and giving out water it may discharge a hydrostatic function, causing the organism to rise or sink in the water.
C. The large central alveole found in the colonies of many Polycyttaria (especially Collosphaerida) and first described in my Monograph (Taf. xxxiv. fig. 1), has since then been observed by Hertwig, Bütschli, and other investigators, and recognised as the "central support of the whole colony, surrounded by a delicate membrane" (compare L. N. 33, p. 31, and L. N. 41, p. 436). In a colony of Trypanosphaera transformata(Pl. 5, fig. 1), which I observed living while in Ceylon in 1881, the membrane of the large central alveole was surrounded by a firm network of sarcoplegma, and could be mechanically isolated from the central jelly-sphere which it enclosed.
D. The pericapsular alveoles, figured in Pl. 4, figs. 2, 3, from a Sphaerozoum, and in Pl. 6, fig. 2, from a Siphonosphaera, were very well preserved in some preparations in the Challenger collection; perhaps their development coincides with the formation of spores, and may be regarded as an encystation.
87. The Extracapsular Fat-Globules.-Fat is probably as widely distributed in the exoplasm as in the endoplasm of the Radiolaria; a considerable proportion of the small, dark, highly refractive granules appear to consist of fat; most likely they are for the most part direct products of metastasis. These widely-spread granules, which are sometimes coloured, and which by their passive motion produce the phenomenon of granular circulation in the exoplasm, are not the only fatty structures in the extracapsulum; larger gbobules sometimes occur. In certain large Collodaria (e.g., Thalassicolla melacapsa, Pl. 1, fig. 5; Thalassophysa sanguinolenta, &c.) radial series of oil-globules are found in the calymma, especially in its proximal portion; in others the central capsule is surrounded by a layer of oil-gbobules (situated in the sarcomatrix). In the PHAEODARIA a part of the phaeodium appears to consist of fat gbobules.
88. The Extracapsular Pigment.-The formation of colouring matters in the extracapsulum is on the whole rare in the Radiolaria, apart from the "yellow cells" (see § 91) and from the peculiar phaeodium of the PHAEODARIA, which will be separately treated of in the next paragraph. Considerable masses of extracapsular pigment, usually black or blue, rarely brown or red, are found only in a few Radiolaria belonging to the first three legions; most often in the SPUMELLARIA. Some large Collodaria, e.g., the common Thalassicolla nucleata and a few other species of this genus (Pl. 1, fig. 4), are characterised by a rich deposit of black or blue pigment in the sarcomatrix and in the proximal portion of the calymma. Brown pigment is deposited in the calymma of many Sphaeroidea and Discoidea, as well as of some NASSELLARIA (Cystidium, Tridictyopus, &c.). In a part of the ACANTHARIA red pigment granules are thickly strewn in the sarcoplegma and pass along the free pseudopodia, as for example in Actinelius purpureus and Acanthostaurus purpurascens. The composition and significance of these extracapsular pigments are not completely known.
On the extracapsular pigment of Thalassicolla nucleata, compare my Monograph, pp. 87, 251. On the red extracapsular pigment-granules of the ACANTHARIA, see L. N. 19, pp. 345, 364, &c.
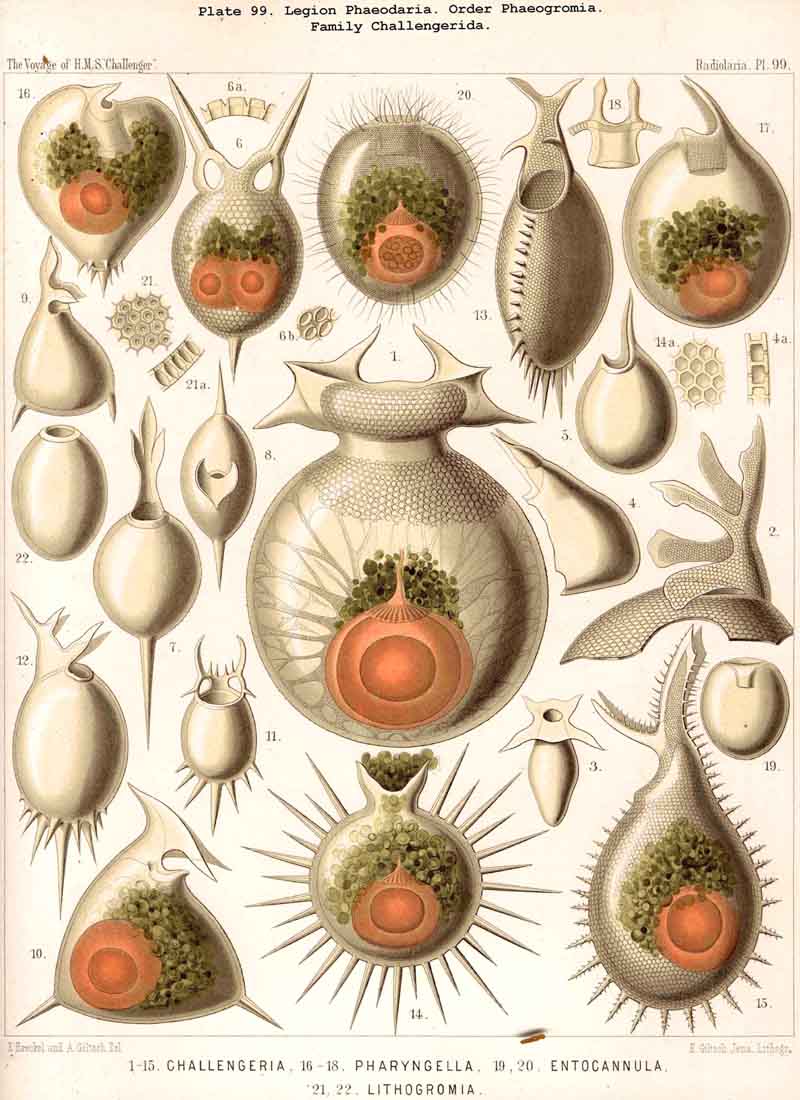
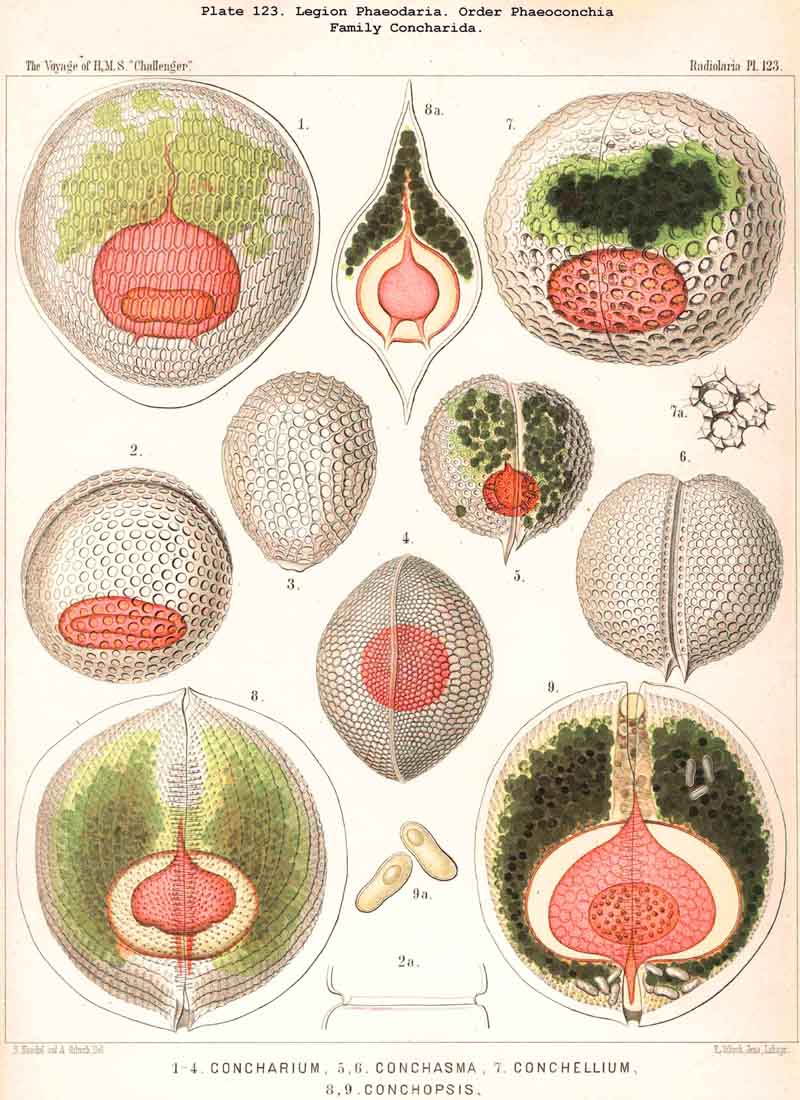
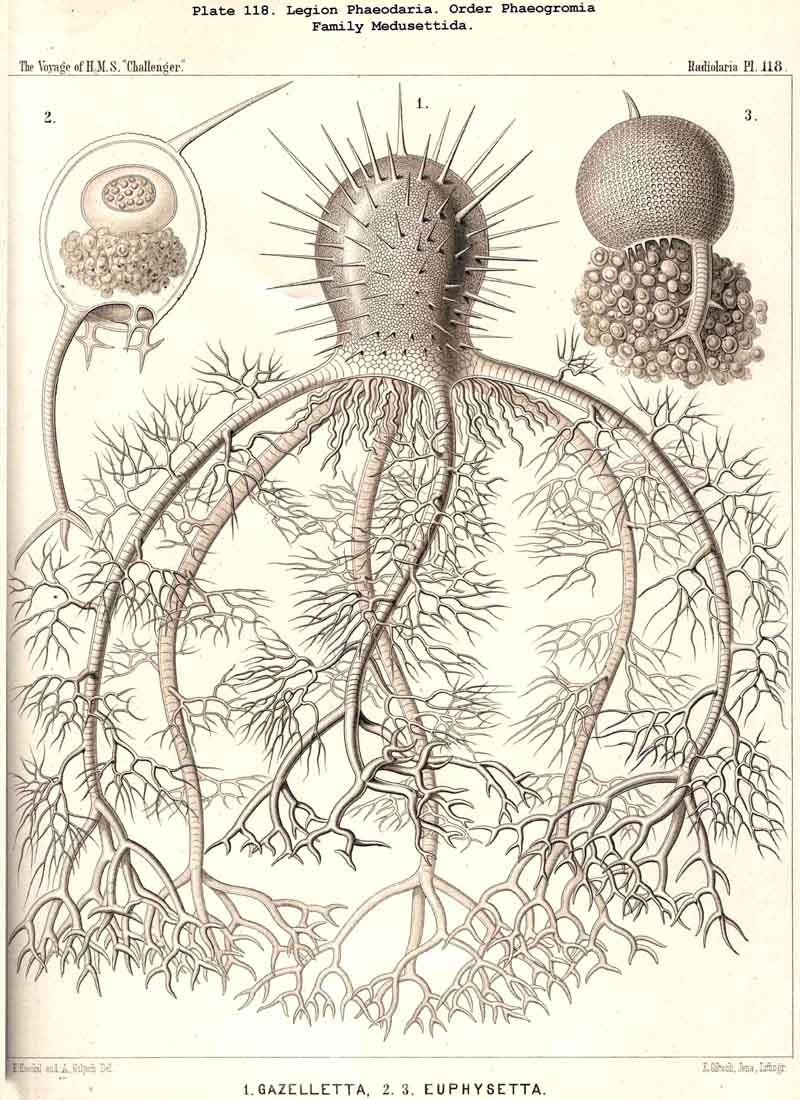
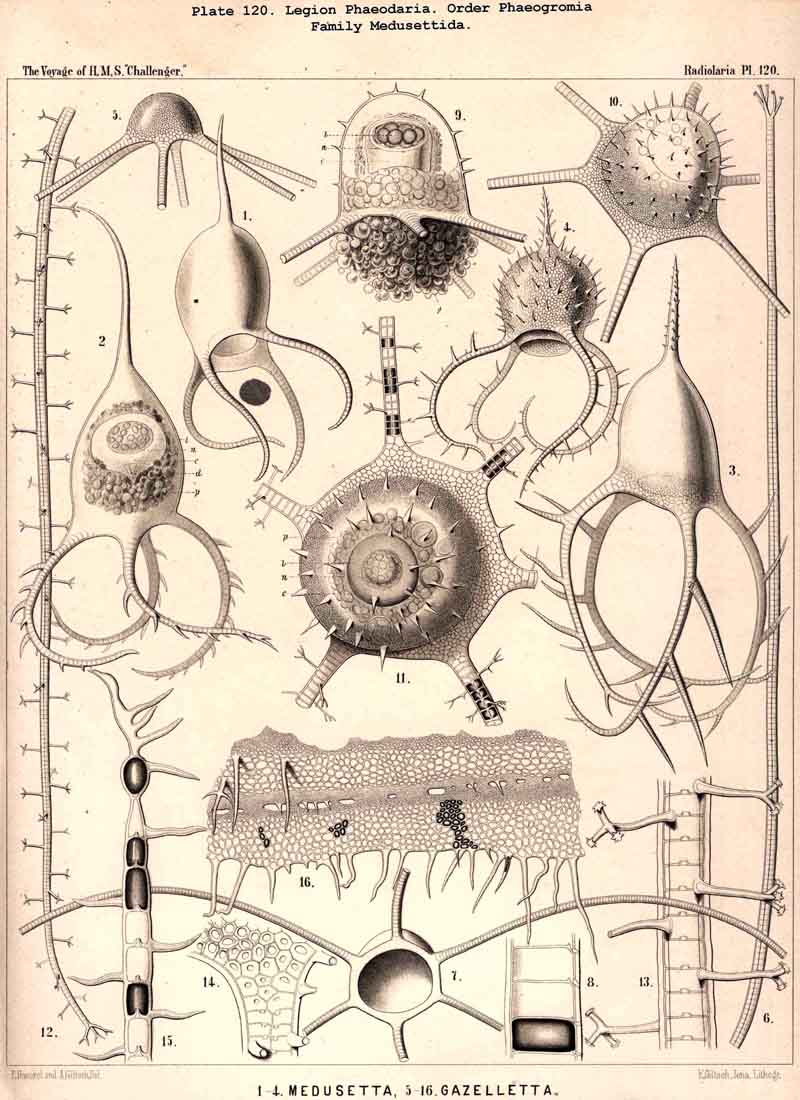
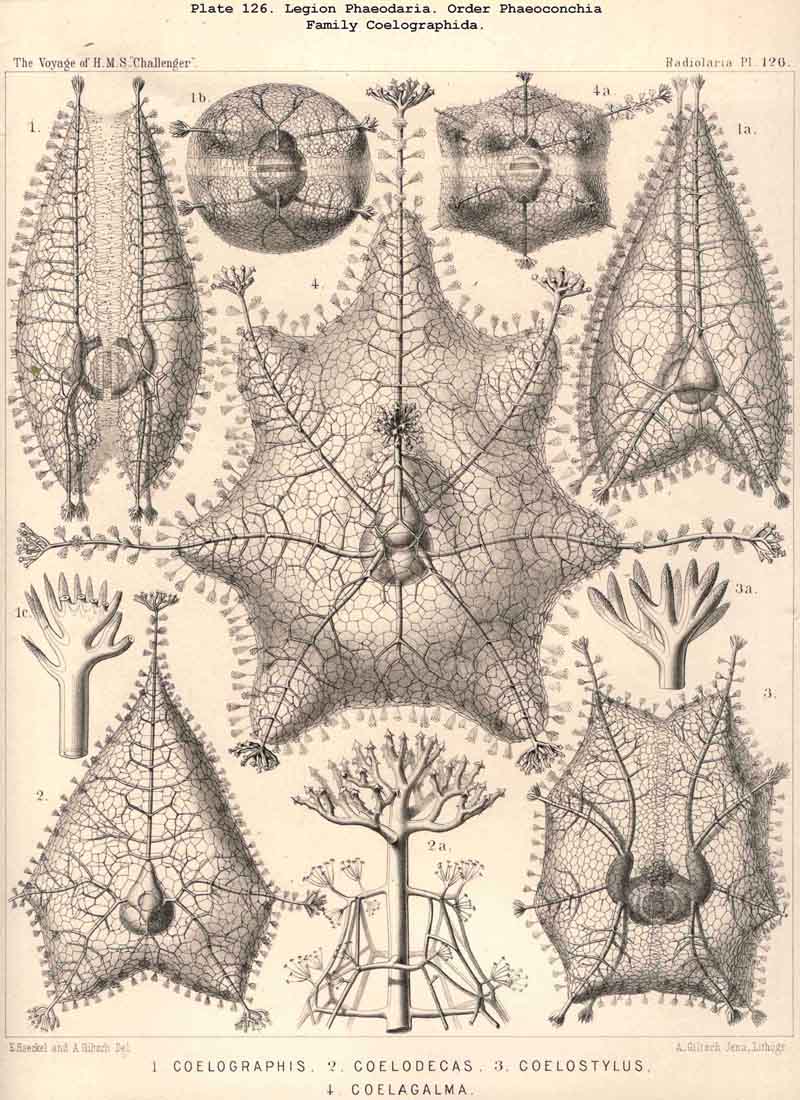
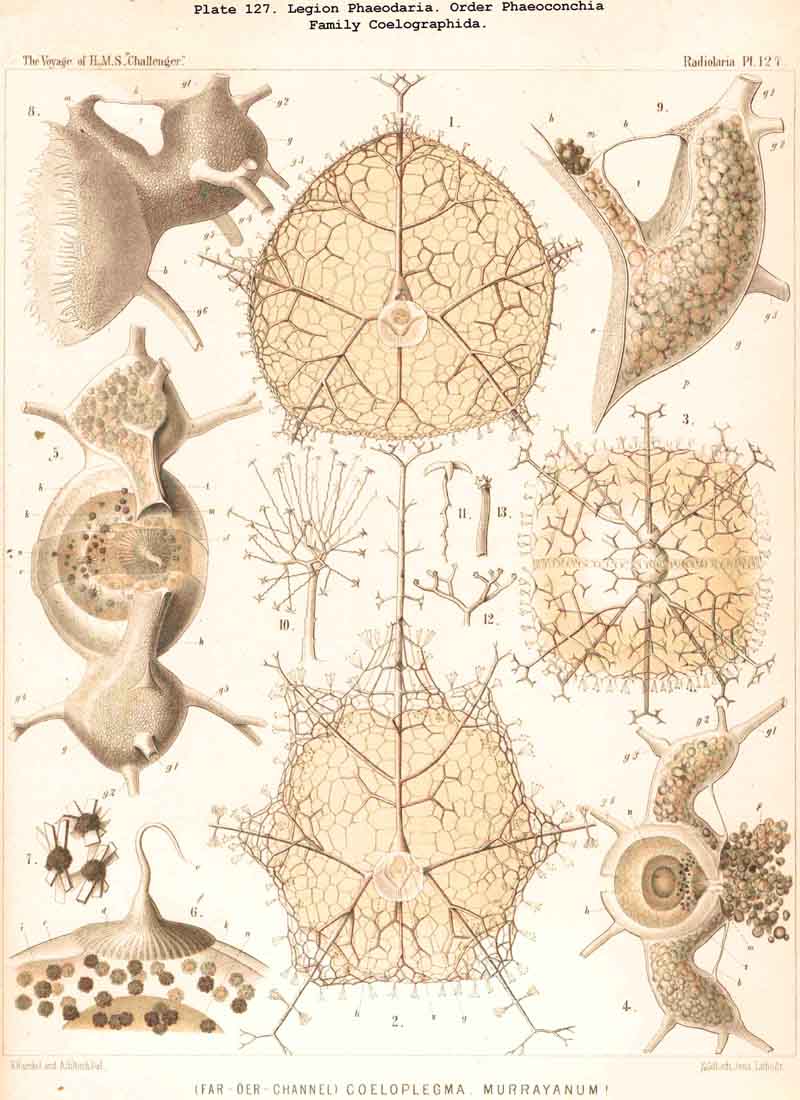
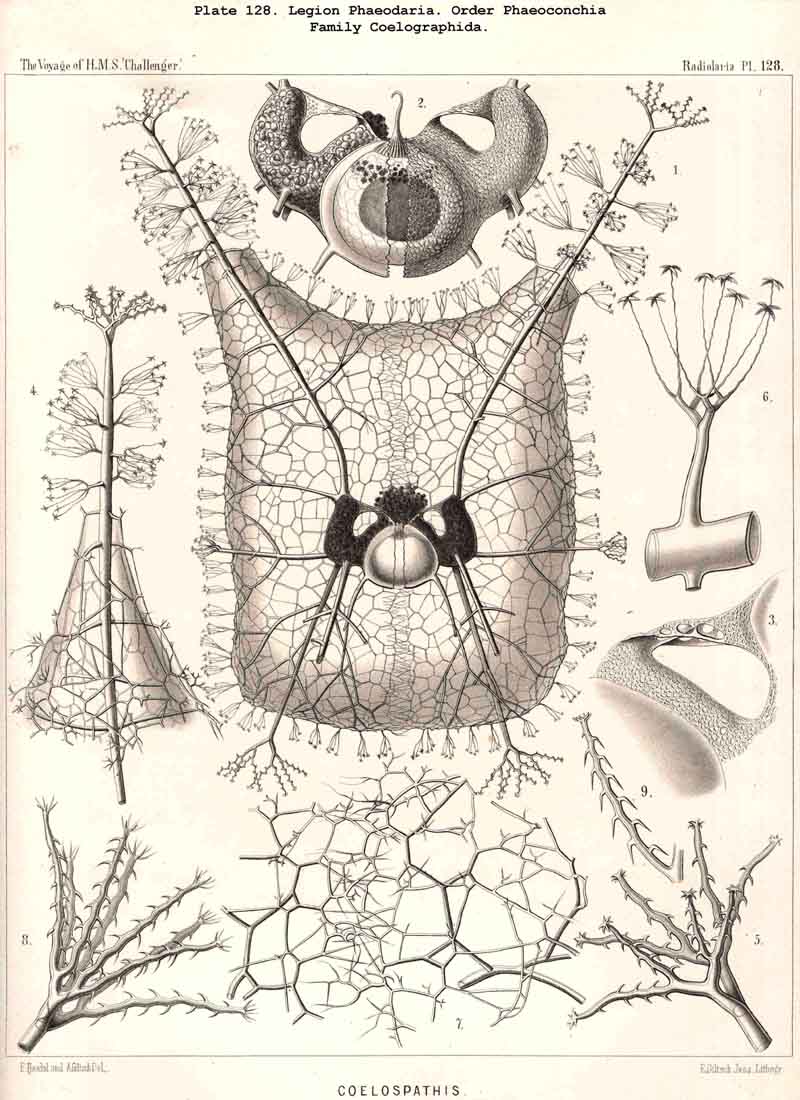
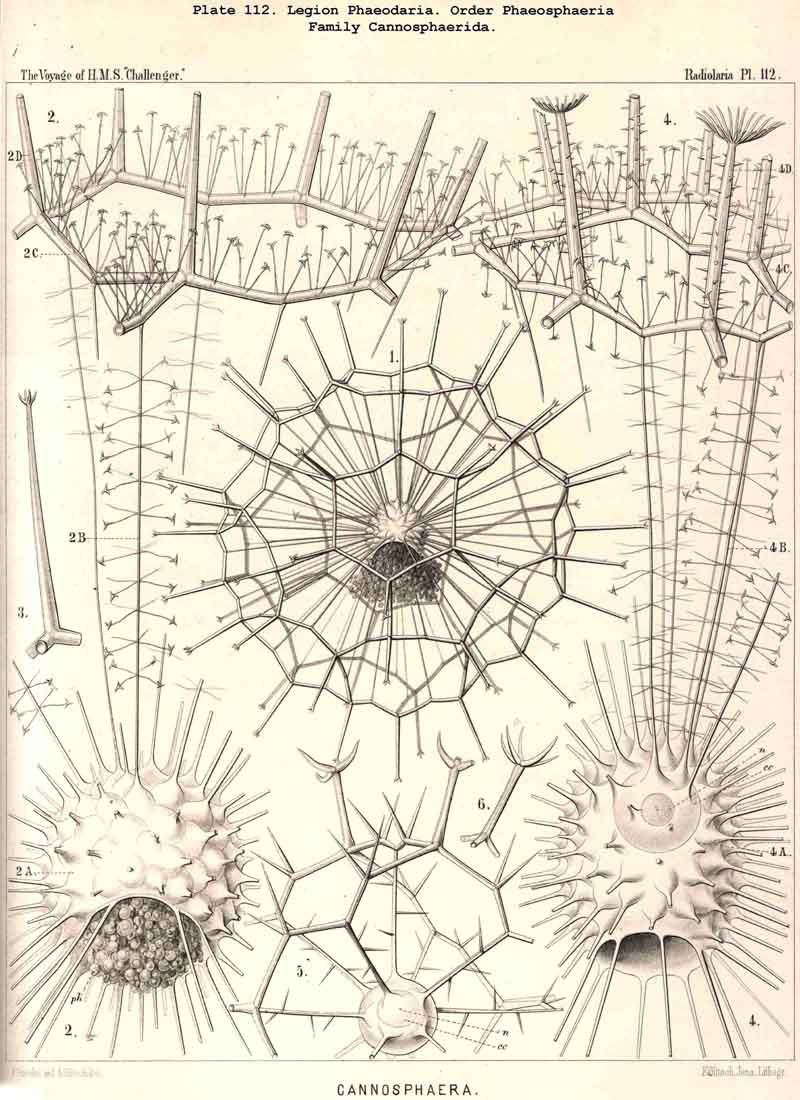
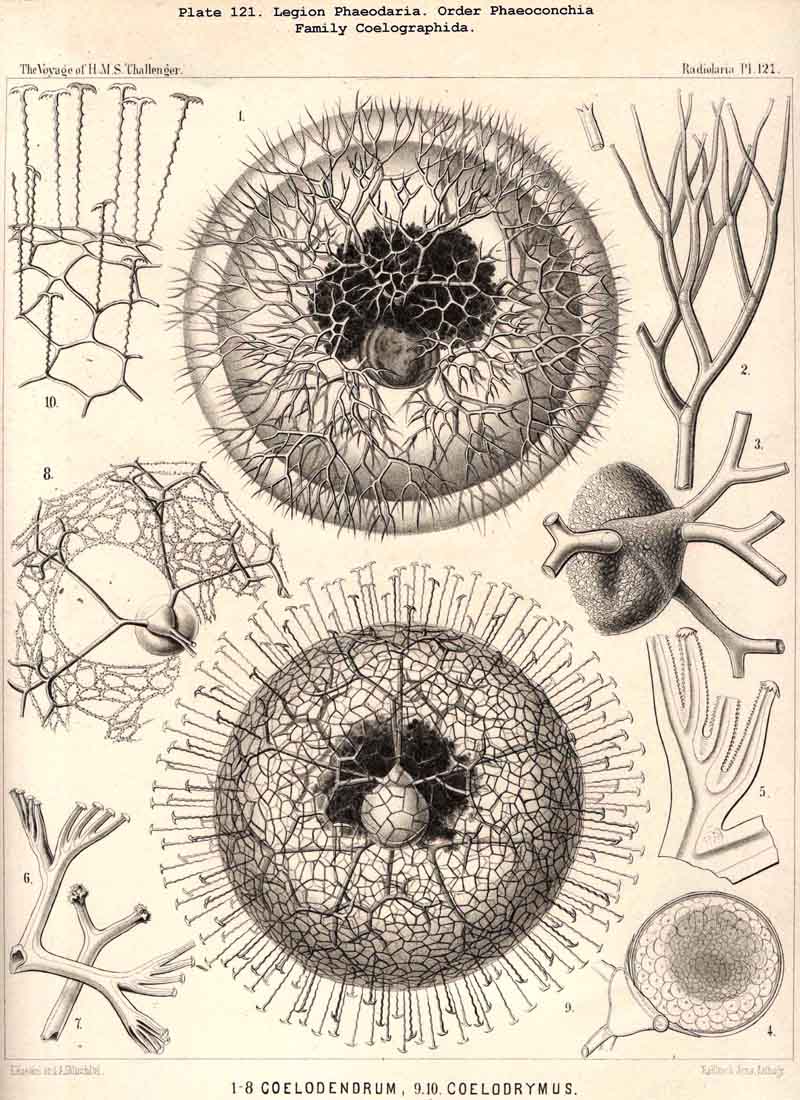
89. The Phaeodium of the Phaeodaria.-The PHAEODARIA, which are distinguished from the other three legions of Radiolaria by the double membrane of the central capsule, and the peculiar structure of the main-opening (astropyle), differ also in other points, the most important of which is the constant presence of a voluminous mass of extracapsular pigment. This possesses a peculiar constitution and special significance, and is not to be confounded with the extracapsular pigment-granules of other Radiolaria (e.g., Thalassicolla), and hence it has been distinguished by the name "Phaeodium," and the individual granules which compose it as "Phaeodella" (see note A). The phaeodium is always excentric in position relatively to the central capsule, of which it surrounds the oral half in the form of a voluminous concavo-convex cap, hiding the astropyle at its basal pole so completely that the latter is rarely visible until the phaeodium has been removed (Pls. 99-104; Pl. 115, fig. 8; Pl. 123, &c.). The central capsule is generally almost completely embedded in the phaeodium, so that only its aboral pole (with the two parapylae in the TRIPYLEA) projects. In the Phaeogromia, in which the lattice-shell possesses a special opening and the central capsule lies excentrically in the aboral portion of its interior, the phaeodium occupies the oral aspect, between the capsule and the aperture (Pls. 99, 100, 118, 119, 120, &c.). In the peculiar family Coelographida (Pls. 126, 127,128) a special receptacle (galea with its rhinocanna) for the phaeodium is developed outside the bivalve shell, within which the central capsule lies. The proboscis, which in all PHAEODARIA arises from the centre of the astropyle, lies in the vertical axis of the phaeodium, and is entirely surrounded by it. The volume of the phaeodium in the majority of the PHAEODARIA may be said to be about as great as that of the central capsule, although in some species it is consider ably larger. Its colour is always dark, usually between green and brown, commonly olive-green or blackish-brown, rarely reddish-brown or black. The phaeodellae or pigment-granules which make up the greater part of the phaeodium (see note B) are irregular in form and unequal in size and show no definite structure; usually they are spherical or ellipsoidal, and exhibit fine parallel striae which run transversely or obliquely (Pl. 101, fig. 3, 6, 10; Pl. 103, fig. 1, &c.). Between the larger granules is usually found a thick dust-like mass of innumerable very small grains. The physiological significance of this peculiar phaeodium is still unknown, but is probably considerable, judging from its large size and especially from its constant topographical relation to the astropyle; the latter consideration would lead to the supposition that it plays an important part in the nutrition and metastasis of the PHAEODARIA (see note C).
A. The phaeodium of Aulacantha, Thalassoplancta, and Coelodendrum was first described in 1862, in my Monograph, as an excentric extracapsular mass of pigment of blackish-brown or olive-green colour (pp. 87, 262, 264, 361, Taf. ii. iii. xxxii.). Since then John Murray, who investigated many living PHAEODARIA during the Challenger expedition, has shown its general distribution in this legion (Proc. Roy. Soc. Lond., vol. xxiv. p. 536, 1876). From the constancy of its presence I gave the legion the name PHAEODARIA in 1879 (L. N. 34).
B. With regard to the special composition of the phaeodium and the constitution of the phaeodellae, see the general description of the PHAEODARIA, pp. 1533-1537.
C. Perhaps the phaeodellae are to some extent symbiontes with the PHAEODARIA; the xanthellae present in most other Radiolaria are absent in this legion.
90. The Extracapsular Xanthellae.- Xanthellae or Zooxanthellae, symbiotic "yellow cells," are very commonly found in the extracapsulum of the Radiolaria, especially in many SPUMELLARIA and NASSELLARIA; whilst in the ACANTHARIA similar yellow cells usually only occur within the central capsule, and in the PHAEODARIA their presence has not been certainly demonstrated. The extracapsular Xanthellae are found most abundantly in the Collodaria, both in the monozootic Thalassicollida and in the polyzootic Sphaerozoida. They occur in smaller numbers in the Sphaerellaria, and in many divisions of the latter they seem to be entirely absent. Also it sometimes happens that, though present in large numbers in some SPUMELLARIA, they are entirely absent in others nearly related to them; indeed, this has also been observed in the case of different individuals of the same species. This fact alone is sufficient to show that the Xanthellae are not an integral part of the Radiolarian organism (as was formerly believed) but parasites or more correctly symbiontes, which live as inhabitants of the calymma. More recent investigations have shown, that besides the yellow pigment-grains they contain starch or an amyloid substance, that is to say, vegetable reserve materials, that their thin envelope contains cellulose, and that their yellow colouring-matter resembles chlorophyll and is related to that of the Diatomaceae ("Diatomin "). Hence they are now generally regarded as unicellular Algae, nearly related to those which occur as symbiontes in other marine animals (Exuviella, &c.). The starch, which they develop with the formation of oxygen, may serve as nutriment to the Radiolaria, while the carbonic acid yielded by the latter is also beneficial to the Xanthellae. The form of the Xanthellae is usually spherical and elliptical, often also sphaeroidal or discoidal. Their diameter is usually between 0.008 and 0.012 mm., rarely more or less. The differences exhibited by Xanthellae which live in different groups of Radiobaria demand further investigation, which will perhaps lead to the establishment of several species of the genus Zooxanthella. At present Zooxanthella extracapsularis, in the calymma of SPUMELLARIA and NASSELLARIA, may be clearly distinguished from Zooxanthella intracapsularis, in the central capsule of the ACANTHARIA.
The "yellow cells" were first described in 1851 by Huxley, in the Collodaria, and afterwards by J. Müller (1858) in many SPUMELLARIA and NASSELLARIA. In my Monograph (1862, pp. 84-87) I gave a detailed account of their structure and increase by division, and laid special emphasis on the fact that they are the only elements in the Radiolarian organism which "are undoubtedly cells in the strict histological sense of the word." Afterwards, in my Beiträge zur Plastiden-Theorie, I showed the constant presence of "starch in the yellow cells of the Radiolaria" (1870, L. N. 21). Shortly afterwards Cienkowski observed that the yellow cells live independently and reproduce themselves after the death of the Radiolaria, and in consequence first put forth the hypothesis that they do not belong to the Radiolarian organism, but that they are unicellular Algae parasitic upon it (1871, L. N. 22). This view was ten years later more fully established by Karl Brandt, and elucidated by comparison with the symbiosis of the gonidia of Algae, and the hyphae of Fungi in the formation of Lichens, which had in the meantime become known (1881, L. N. 38). Brandt gave this unicellular yellow Alga the name Zooxanthella nutricola, and afterwards gave fuller details regarding its remarkable vital relations (L. N. 39), Patrick Geddes, who named it Philozoom, supplemented this account and showed experimentally that it gives off oxygen under the influence of sun-light (1882, L. N. 42, 43). In consequence of this there is no doubt that all Xanthellae (the Zooxanthella extracapsularis of SPUMELLARIA and NASSELLARIA, and the Zooxanthella intracapsularis of the ACANTHARIA, and possibly also the Zooxanthella phaeodaris of the PHAEODARIA) do not originally belong to the Radiolarian organism, as was believed up to the time of Cienkowski, but, penetrate actively into it from without, or are taken in passively by means of the pseudopodia. In any case their symbiosis, when they are associated with the Radiolarian cell in large numbers, may be of great advantage to both parties, since the metastasis of the Xanthella is vegetable, that of the Radiolarian animal in character. In any case their symbiosis is to a large extent accidental, by no means as necessary as in the case of the Lichens. See on these points in addition to Brandt and Geddes (loc. cit.) also Geza Enz, Das Consortial-Verhältniss von Algen und Thieren, Biol. Centralbl., Bd. ii. No. 15, 1883, Oskar Hertwig, Die Symbiose oder das Genossenschaftsleben im Thierreich, Jena, 1883, and Bütschli, Die Radiolarien, in Bronn's Klass. u. Ord. d. Thierreichs, 1882 (L. N. 41, pp. 456-462).
91. The Exoplasm or Extracapsular Protoplasm.-The extracapsular protoplasm, which may be shortly termed the "exoplasm" (or ectosarc), is primitively in all Radiolaria (and especially in their earliest development stages) the only important constituent of the extracapsulum, besides the calymma. Although the extracapsular and intracapsular protoplasm of the Radiolaria are everywhere in direct communication, and although the openings in the membrane of the central capsule bring about an interchange between them, still the two portions of sarcode show certain constant and characteristic differences, which are due to the physiological division of labour between the central and peripheral parts of the body and their corresponding morphological differentiation. The extracapsular, like the intracapsular, protoplasm is originally homogeneous, but may afterwards become differentiated in various ways, producing the special constituents of the extracapsulum. Such "external protoplasmic products" are vacuoles, pigment-bodies, &c. More important, however, are the topographically different sections into which the exoplasm may be divided according to its relations to the central capsule and the calymma. In this respect the following parts may be generally distinguished- ( 1) the Sarcomatrix, or fundamental layer of the exoplasm, which surrounds the central capsule as a continuous sheath of sarcode and separates it from the calymma; (2) the Sarcoplegma, an irregular network of the exoplasm, which spreads throughout the gelatinous material of the calymma; (3) the Sarcodictyum or network of sarcode on the outer surface of the calymma; and (4) the Pseudopodia, which project outwards from the latter and radiate into the water.
92. The Sarcomatrix.-The sarcomatrix, being "the fundamental layer of the pseudopodia" (or "matrix of the exoplasm "), constitutes the proximal innermost section of the extracapsular sarcode, and in all Radiolaria forms a thin continuous mucous layer, which covers the whole outer surface of the central capsule and separates it from the surrounding calymma (see note A, below). The sarcomatrix communicates internally through the openings of the central capsule with the endoplasm, whilst externally the pseudopodia or mucous threads arise from it, which by their union form the sarcoplegma. The sarcomatrix is only interrupted in the SPUMELLARIA and ACANTHARIA by those parts of the skeleton which perforate the membrane of the central capsule. In all NASSELLARIA and PHAEODARIA, as in the Collodaria, it appears as a perfectly continuous sarcode-envelope of the central capsule. Its thickness is variable; in general it is most strongly developed in the SPUMELLARIA and PHAEODARIA, less so in the NASSELLARIA, and is thinnest in the ACANTHARIA. The thickness seems, however, to vary even in one and the same individual, the difference depending partly upon the different stages of development and partly upon nutritional conditions. After abundant inception of nutriment the thin protoplasmic layer of the matrix is thickened and turbid, rich in granules and irregular masses, which are probably due to enclosed but only half-digested food; xanthellae also, as well as foreign bodies taken up with the nutriment, such as frustules of Diatoms and shells of smaller Radiolaria, and of pelagic infusoria, larvae, &c., are often, especially in large individuals, aggregated in consider able quantities in the matrix. After long fasting, on the contrary, this is poor in these enclosed bodies and in granules; it then forms a thin colourless more or less hyaline mucous coating to the central capsule. From a physiological standpoint the sarcomatrix is to be regarded as the central organ of the extracapsulum, and as of preeminent significance. Probably it is not only the most important organ for the nutrition of the Radiolaria (especially for digestion and assimilation in particular), but perhaps is also the central organ of perception. On the other hand the sarcomatrix belongs to those components of the Radiolarian organism which take no part in the formation of the skeleton.
A. The sarcomatrix was first described in my Monograph in 1862 (p. 110) as the "Mutterboden der Pseudopodien," possessing a pre-eminent physiological importance. Compare also my paper on the sarcode elements of the Rhizopoda (Zeitschr. f. wiss. Zool., Bd. xv. p. 342, 1865).
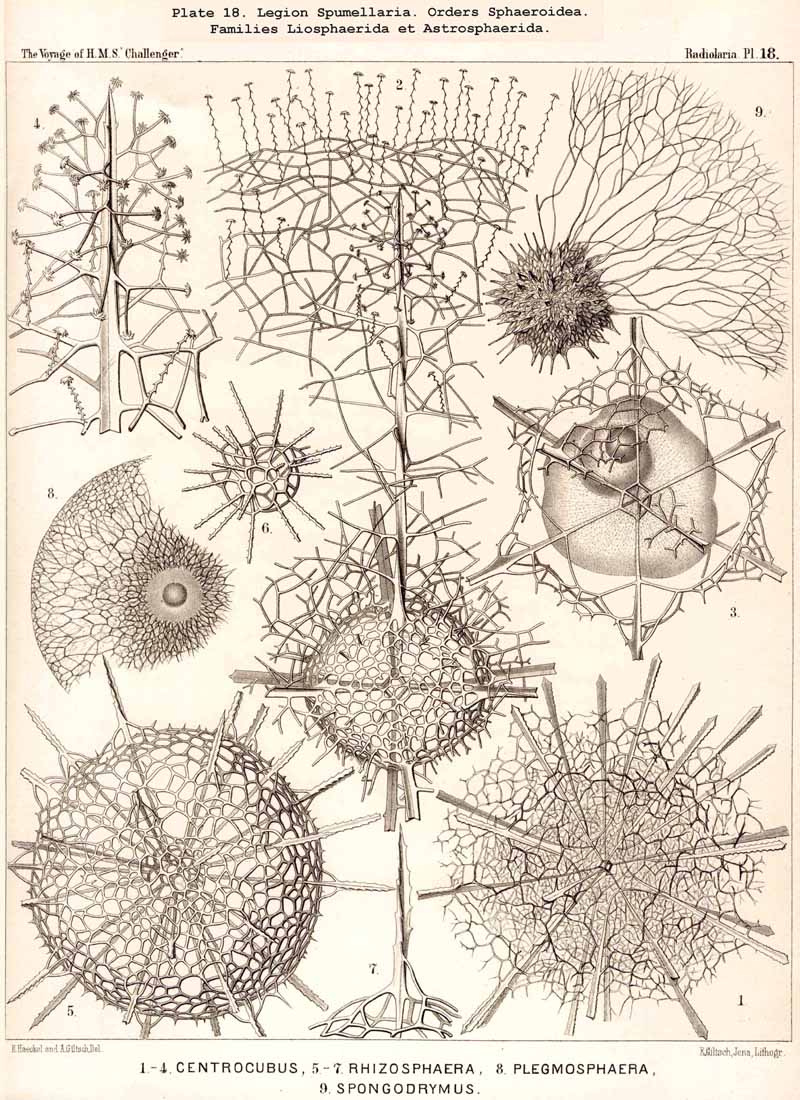
93. The Sarcoplegma.-By the name sarcoplegma, as distinguishcd from the remaining extracapsular sarcode, is understood the intracalymmar web of exoplasm or "ectosarcode network," which ramifies within the gelatinous mass of the calymma. Internally it is in direct connection with the continuous sheath (sarcomatrix), which encloses the central capsule, whilst externally it is in contact with the superficial sarcode network (sarcodictyum) which surrounds the calymma. The configuration of this exoplasmic web, which penetrates the jelly-veil in all directions, is exceedingly variable; in most Radiolaria it is extremely irregular in form, like the protoplasmic network in the ground-substance of many kinds of connective tissue. In some groups, however, it assumes a rather regular shape which it appears to retain (e.g., in many ACANTHARIA). It must be assumed also that in those instances where the consistency of the calymma approaches that of cartilage, the tracks of the exoplasmic threads remain constant, but accurate observations are wanting as to how far the configuration of the sarcoplegma is constant or variable in the different groups, as well as regarding its peculiar behaviour in those Radiolaria whose calymma is characterised by the formation of vacuoles or alveoles (see § 86). Usually it envelops the larger alveoles in the form of a reticulate veil. In many Collodaria the exoplasm is aggregated at certain points of the intracalymmar web, so that large balls or amoeboid bodies appear to be distributed between the alveoles, e.g., in Thalassophysa pelagica and Thalassicolla melacapsa (Pl. 1, figs. 4, 5). The sarcoplegma is metamorphosed directly into silex in the Radiolaria spongiosa, or those genera which possess a spongy cortical skeleton, and were formerly known as Spongurida; to this category belong the Spongosphaerida (Pl. 18) and Spongodiscida (Pl. 47) as well as certain NASSELLARIA and PHAEODARIA. The single siliceous spicules, which are irregularly interwoven to form the spongy web, are to be regarded as the silicified threads of the intracalymmar sarcode network. From a physiological point of view the sarcoplegma is of importance both for the nutrition and motion of the Radiolaria, since it brings the sarcomatrix and the sarcodictyum, with the pseudopodia which radiate from it, into direct communication.
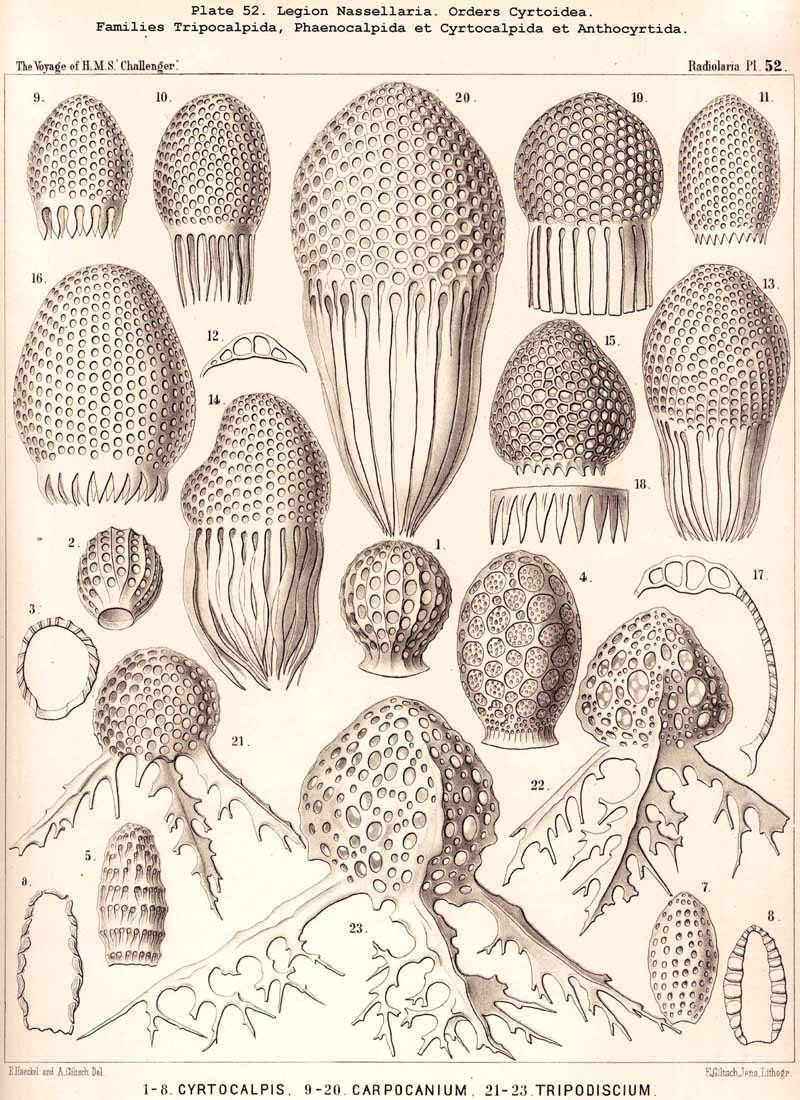
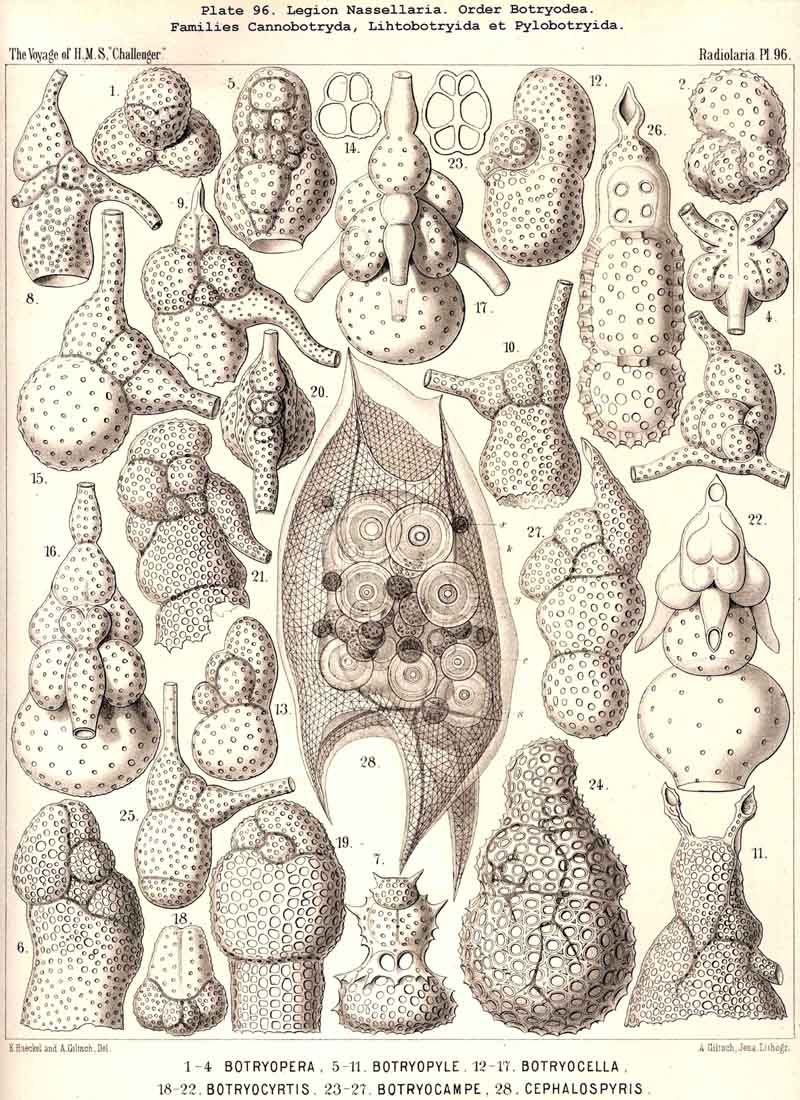
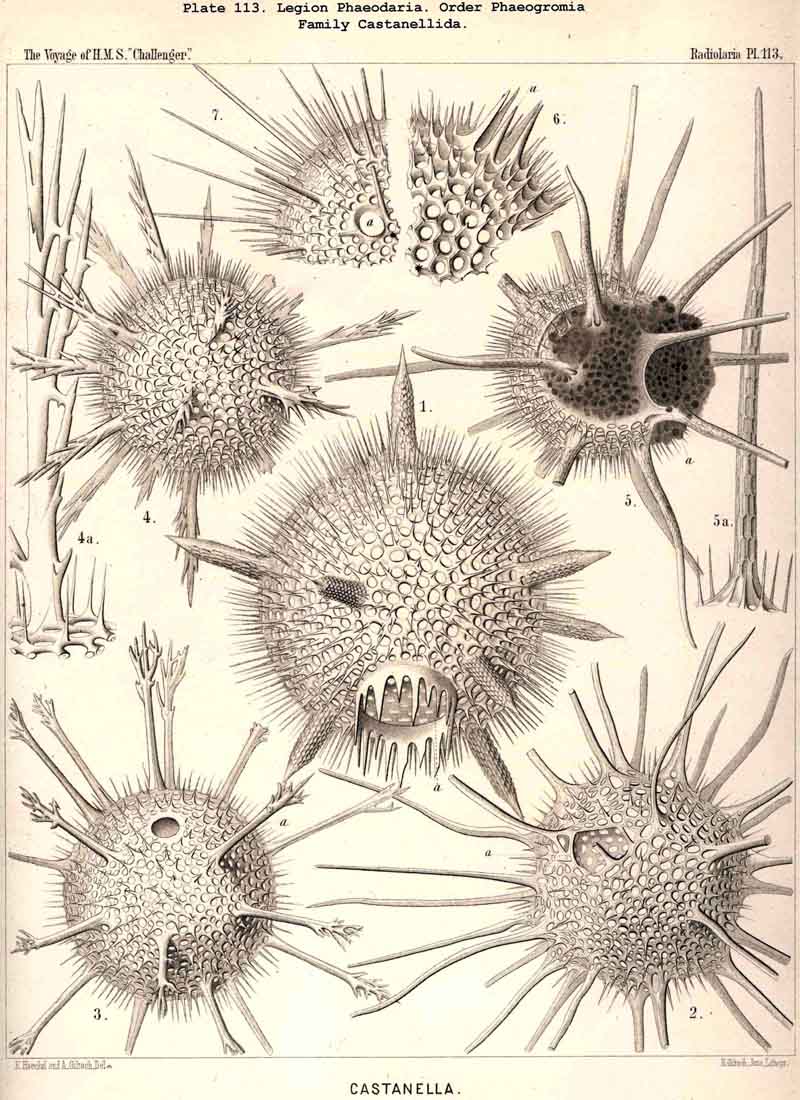
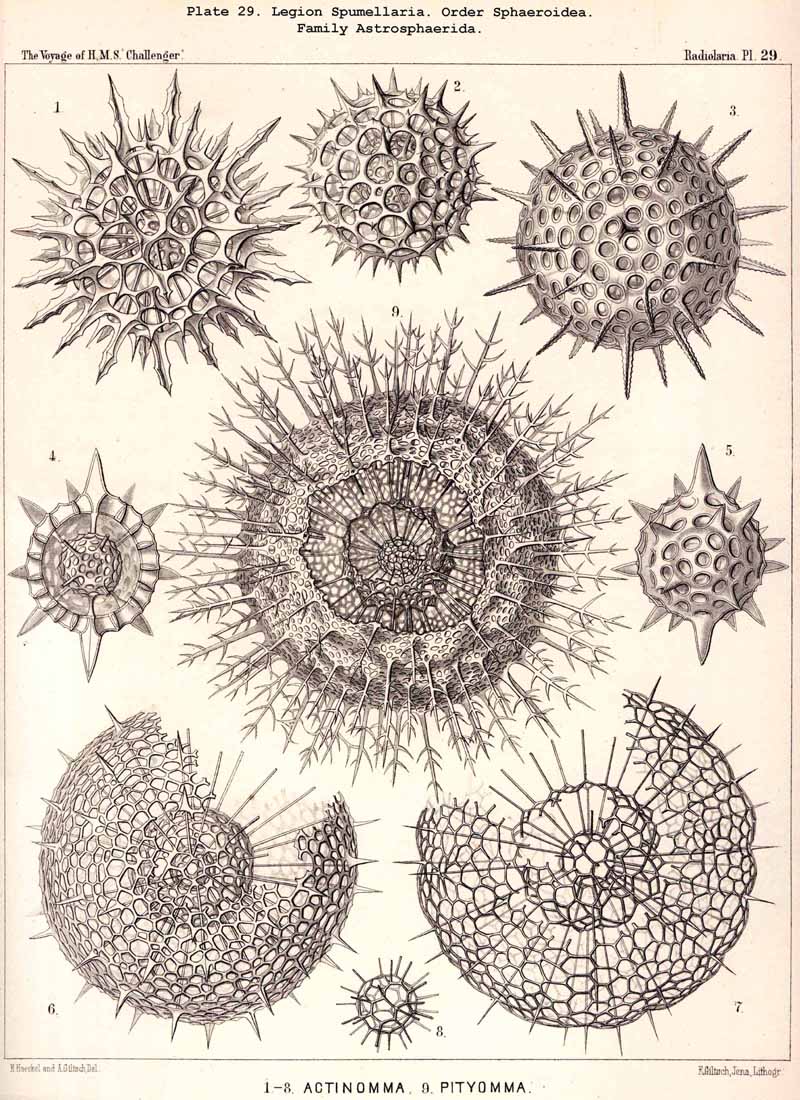
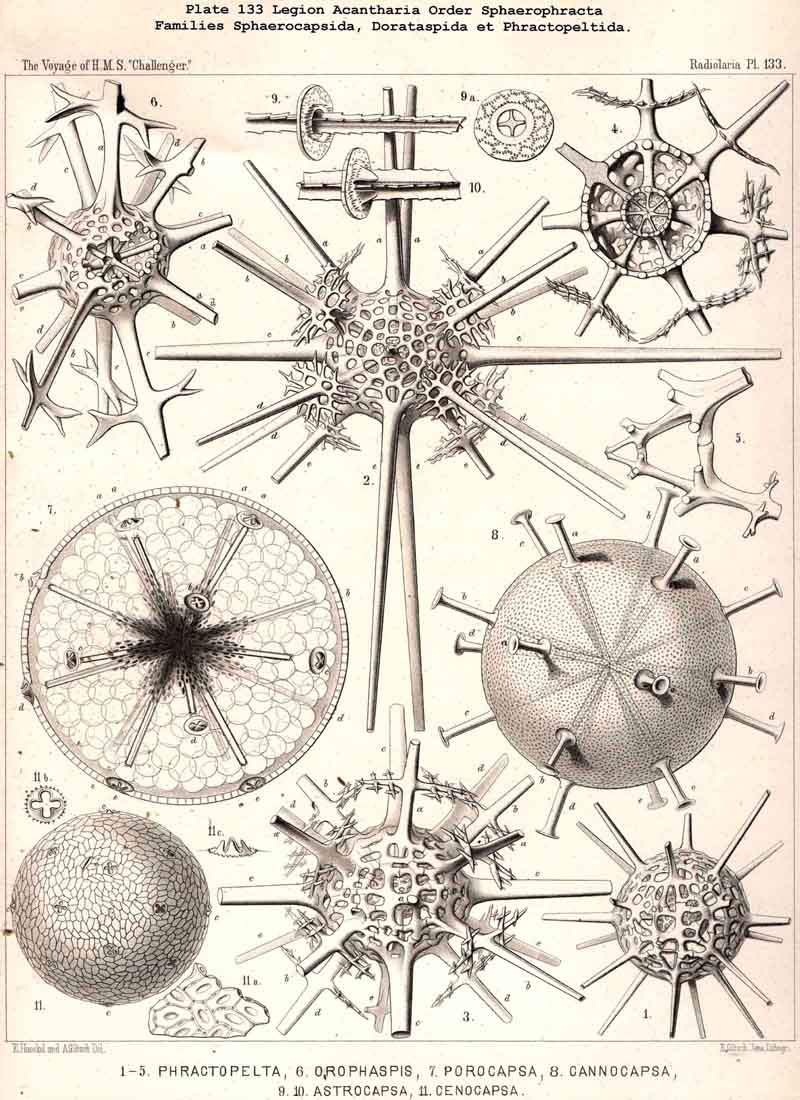
94. The Sarcodictyum.-The sarcodictyum may be defined as the extracalymmar network of exoplasm, and is a reticular covering which lies upon the outer surface of the gelatinous calymma. Internally, the sarcodictyum is in direct communication with the sarcoplegma, or the web of exoplasmic threads which ramifies in the gelatinous substance of the calymma; externally, on the other hand, the pseudopodia radiate freely from it; thus its relation to these is similar to that which the sarcomatrix bears to the roots of the sarcoplegma. Relations similar to those which have led to the separation of the primary from the secondary calymma, induce us to distinguish also a primary and secondary sarcodictyum. The original or primary sarcodictyum ramifies over the surface of the original or primary calymma, and like this is of pre-eminent importance in the formation of the primary lattice-shell; if we regard the surface of the primary calymma as the indispensable foundation for the deposition of this latter, then the primary sarcodictyum furnishes the material from which it is developed: silex in the SPUMELLARIA and NASSELLARIA, a silicate of carbon in the PHAEODARIA, and acanthin in the ACANTHARIA. It may indeed be said that the primary lattice-shell of the Radiolaria arises by a direct chemical metamorphosis of the primary sarcodictyum, by a chemical precipitation of the dissolved skeletal material (silex, silicate, or acanthin), which was stored up in the exoplasm of the sarcodictyum. Hence a deduction from the special conformation of the former to that of the latter is permissible. The particular form of the primary lattice-sphere with its regular or irregular meshes is due to the corre sponding form of the primary sarcodictyum; both regular and irregular forms of this commonly occurring. The form of the regular sarcodictyum with circular or regular polygonal, usually hexagonal, meshes is constantly maintained during the formation of the regular lattice-shells (e.g., Pl. 12, figs. 5-10; Pl. 52, figs. 8-20; Pl. 96, figs. 2-6; Pl. 113, figs. 1-6). The form of the irregular sarcodictyum, on the other hand, with irregular polygonal or roundish meshes, persists during the development of the irregular lattice-shells (e.g., Pls. 29, 70, 97, 106). All this is true also of the secondary sarcodictyum, or the exoplasmic network which ramifies over the surface of the secondary calymma. The secondary lattice-shells, which are deposited on the surface of the latter, retain the configuration of the secondary sarcodi ctyum, by the chemical metamorphosis of which they have originated; this is the case in many SPUMELLARIA which develop several concentric lattice-shells (Pl. 29), in some NASSELLARIA (Pl. 54, fig. 5), in the Phractopeltida among the ACAINTHARIA (Pl. 133), and in the double shelled PHAEODARIA, Cannosphaerida, and part of the Coelodendrida and Coelographida (Pls. 112, 121, 128). In those Radiolaria which form no lattice-shell whatever, the conformation of the sarcodictyum is usually irregular, with meshes of irregular form and unequal size; sometimes, however, they seem to be very regular, as in many Acanthometra (Pl. 129, fig. 4).
95. The Pseudopodia.-On the whole the pseudopodia or thread-like processes of the exoplasm exhibit in the Radiolaria the same characteristic peculiarities as in all true Rhizopoda; they are usually very numerous, long and thin, flexible and sensitive filaments of sarcode, which show the peculiar phenomena of granular movement. Their physiological significance is in several respects very great, for they serve as active organs for the inception of nutriment, for locomotion, sensation, and the formation of the skeleton (see note A, below). The presence of a calymma, however, which distinguishes the Radiolaria from the other Rhizopoda, brings about certain modifications in the behaviour of the pseudopodia. If in general all the threads, which arise from the sarcomatrix or fundamental layer and radiate outwards, be called "pseudopodia," then that part of them which is included in the gelatinous substance of the calymma and forms the sarcoplegma may be termed the "collopodia" (or intracalymmar pseudopodia), and the remaining portion, which passes outwards from the sarcodictyum freely into the water, may be described as "astropodia" (or extracalymmar pseudopodia). In many Radiolaria these two portions present some differences in morphological and physiological respects, and certain distinctions are probably generally present (see note B). Apart from this universal differentiation in the different groups of the Radiolaria, specially modified forms of pseudopodia may be recognised as the axopodia and myxopodia of the ACANTHARIA (see § 95, A), and the sarcode-flagellum of certain SPUMELLARIA (see note C).
A. The pseudopodia of the Radiolaria have been so fully described in my Monograph, in 1862, both morphologically and physiologically, that I need only refer to the account there given (pp. 89-127); for supplementary observations see II. Hertwig (1879, L. N. 33, p. 117) and Bütschli (1882, L. N. 41, pp. 437-445).
B. The Astropodia, or free radiating pseudopodia, are in many Radiolaria more or less clearly distinguishable from the collopodia, which form the sarcoplegma within the calymma; how far these distinctions depend upon a permanent differentiation (especially in the ACANTHARIA and PHAEODARIA) needs further investigation.
C. The sarcode-flagellum (perhaps better termed axoflagellum) was first described in my Monograph (1862, p. 115) in the case of various Discoidea (Taf. xxviii. figs. 5,8; Taf. xxx. fig. 1). Hertwig has given a substantially similar account of the organ in some other Discoidea (L. N. 33, p. 67, Taf. vi. figs. 10, 11); probably this peculiar structure is confined to the order Discoidea among the SPUMELLARIA, but is widely distributed within its limits. The axoflagellum is a thick cylindrical thread of sarcode, finely striated and pointed towards its free end. It always lies in the equatorial plane of the discoidal body, and always unpaired in one of its axes; in the triradiate Discoidea it is in the axis of the unpaired principal arm and opposite to it (Pl. 43, fig. 15). In the Ommatodiscida (p. 500, Pl 48, figs. 8, 19, 20) the axoflagellum probably passes out through the peculiar marginal ostium of the shell. Perhaps it is always connected with the central nucleus by intracapsular axial fibres, and is to be regarded as a specially differentiated bundle of pseudopodia (or axopodia?).
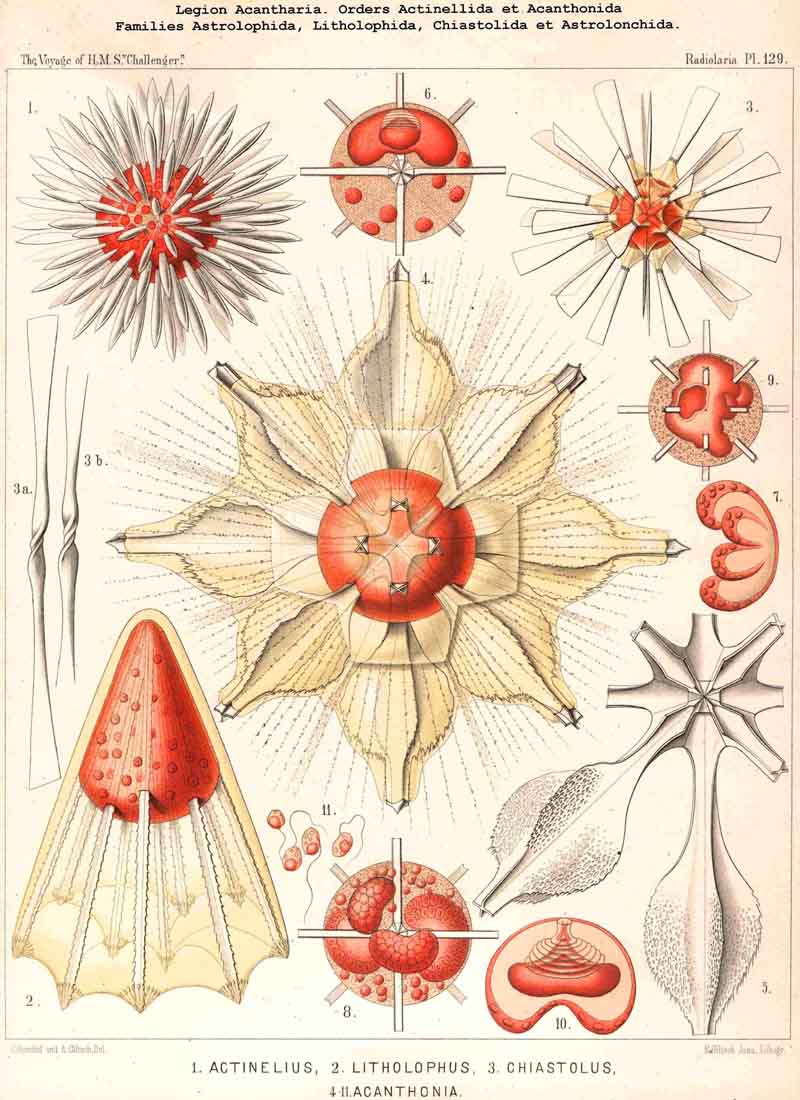
95A. The Myxopodia and Axopodia.-The two forms of pseudopodia which we distinguish as myxopodia and axopodia differ markedly from each other both morphobogically and physiologically. The myxopodia, or ordinary free pseudopodia, which are found in large numbers in all Radiolaria, and constitute their most important peripheral organs, are simple homogeneous exoplasmic threads, which arise from the sarcodictyum or extracalymmar sarcode network, and radiate freely into the water; here they may branch and combine by anastomosis to form a changeable network, but they never contain an axial thread. The axopodia, on the other hand, are differentiated pseudopodia, which consist of a firm radial thread, and a soft covering of exoplasm ; they penetrate the whole calymma in a radial direction and project freely from its surface, and generally (if not always) they are produced inwards to the middle of the central capsule, perforating its membrane; their proximal end is lost in a dark central heap of granules. Such axopodia are at present known with certainty only in the ACANTHARIA, where they are widely, and perhaps universally, distributed. Their develop ment in this legion probably stands in direct causal relation to the peculiar structure of the central capsule and the centrogenous formation of the skeleton. Since the radial skeletal rods of the Acanthometra possess originally a thin coating of protoplasm, it may be said that the centrogenous axopodia of this group became differentiated in two ways, the firm axial threads of one section remaining very thin and covered by protoplasm, whilst those of the other section became metamorphosed into radial bars of acanthin. This hypothesis acquires more probability from the regular distribution and arrangement of the axopodia in the ACANTHARIA; they usually stand at fixed intervals between the radial bars, singly or in groups; sometimes their number seems to be not greater than that of the bars, whilst in other cases a circlet or group of axopodia corresponds to each radial bar. Perhaps their fine axial thread consists of acanthin. At all events the axopodia are constant organs (probably sensory, like the "palpocils") and not retractile like the movable myxopodia.
The axial threads in the pseudopodia of the Acanthometra were first discovered by R. Hertwig, who accurately described their peculiar structure and arrangement (L. N. 33, pp. 16, 117).
96. The Myophriscs of the Acanthometra.- The Acanthometra are characterised by a very peculiar differentiation of the exoplasm, namely, by the formation of myophriscs or contractile threads from the sarcodictyum. In most (and perhaps in all) ACANTHARIA of this order each radial bar is surrounded by a circlet of such contractile threads, which was first described as a "ciliary corona" (see note A, below). The number of contractile threads in each circlet usually amounts to from ten to twenty, rarely being more than thirty and less than eight ; it often appears to be constant in the individual species (see note B). In the living state the myophriscs are long, thin filaments, the pointed distal end of which is inserted into the radial bar, whilst the thicker proximal end is attached to the surface of the calymma, which is elevated round the base of each rod into the form of a gelatinous cone or skeletal sheath (see note C). Probably the myophriscs lie on the outer surface of the apical portion of this gelatinous cone, and are hence to be regarded as exoplasmic threads differentiated from the sarcodictyum. Sometimes, however (as in Acanthochiasma), they fuse into a contractile membrane and form the envelope of a cone, whose interior is occupied by a gelatinous papilla of the calymma. On mechanical irritation the myophriscs contract rapidly and suddenly, like muscle-fibrillae, becoming at the same time thicker, and hence are very different from pseudopodia. Their distal point of insertion being fixed to the firm acanthin rod, they raise by their contraction the skeletal sheath, to which their bases are attached or in the surface of which they lie. The result of their contraction is therefore a distention and increase in volume of the calymma, with which is no doubt connected an inception of water into the gelatinous mass, and hence a diminution in its specific gravity. Probably the Acanthometra contract their myophriscs voluntarily when they wish to rise in the water ; when these relax the calymma collapses owing to its elasticity, water is then expelled and the specific gravity increases. From a physiological point of view, then, the myophniscs are to be regarded as a hydrostatic apparatus, morphologically as myo phanes or muscular fibnillae, such as also occur in the intracapsular protoplasm (see §§ 77-80). On more violent irritation and after the death of the Acanthometra the myophriscs separate from the radial bars and remain attached to the distal ends of the conical gelatinous sheaths as free "ciliary coronas." At the same time, they melt into short, thick, hyaline rods, the so-called "gelatinous cilia." The myophriscs are found only in the order Acanthometra, and are wanting in the Acanthophracta, as well as in the other three legions of Radiolaria.
A. The "ciliary coronas" on the skeletal rods of dead Acanthometra were first described by the discoverer of this order, Johannes Müller, and referred to as "the stumps of the contracted, thickened threads" (L. N. 12, p. 11, Taf. xi.).
B. The "number of thc gelatinous cilia." I found constant in certain species of Acanthometra, and stated in my Monograph (L. N. 16, p. 115) "that here is to be found the first differentiation of the diffuse sarcode into definite organs of regular definite number, size, and position, which deserve the name tentacles rather than pseudopodia."
C. The nature of the myophriscs as fibrillae allied to muscles was first discovered by R. Hertwig, who described them as "structures of peculiar nature," under the name of "contractile threads," and pointed out in detail their histological and physiological peculiarities (L. N. 33, pp. 16-19, Taf. i).
97. The Exoplasm of the Peripylea.-The extracapsular protoplasm of the SPUMELLARIA or PERIPYLEA is in communication with the intracapsular sarcode by the innumerable fine pores of the capsule-membrane, and like these pores is evenly distributed over the whole surface. The sarcomatrix which immediately surrounds the central capsule is moderately strong, and sends out innumerable long, thin pseudopodia, which probably correspond to the pores of the membrane. Their number is markedly greater in the SPUMELLARIA than in the other three legions. The ramifications and communications which the radiating fibres of the sarcomatrix undergo within the calymma, apparently present the most manifold variations, so that the sarcoplegma or intracalymmar network thus formed has very diverse forms. On the surface of the calymma the exoplasmic threads constitute a variously disposed sarcodictyum, a regular or irregular exoplasmic network, by the silicification of which a primary lattice-shell arises in the majority of the SPUMELLARIA. The free ends of the pseudopodia, which arise from this extracalymmar network and radiate out into the water, appear in most SPUMELLARIA to be relatively short, but exceedingly numerous. Specially modified pseudopodia and axial threads in particular do not seem to occur in this legion. Perhaps, however, among the latter may be reckoned the remarkable pseudopodia which combine to form the sarcode flagellum in many Discoidea (and perhaps in other SPUMELLARIA). This axoflagellum is a particularly strong thread of sarcode, arising from a definite point in the central capsule ; it is cylindrical or slenderly conical in form, much longer, stronger, and more contractile than the ordinary pseudopodia ; it contracts in a serpentine fashion on mechanical irritation and seems to originate by the fusion of a bundle of pseudopodia (compare § 95, C).
98. The Exoplasm of the Actipylea.-The extracapsular protoplasm of the ACANTHARIA or ACTIPYLEA differs in several important respects from that of other Radiolaria, and appears to undergo more significant differentiations than that of the three other legions. Since the pores in the wall of the central capsule are not distributed evenly and at equal intervals over its whole surface (as in the PERIPYLEA), but rather exhibit a regular disposition in groups at unequal intervals, the number of projecting pseudopodia is much less and the law of their arrangement different from that which obtains in the PERIPYLEA (§ 58). In many and probably in all ACANTHARIA they are divided into two groups, those which arise from the centre of the capsule and possess firm axial threads, and those which have not these characters (compare § 95, A). The axopodia, or stiff pseudopodia with axial threads, arise from the centre of the capsule, are present in much smaller numbers than the soft and flexible myxopodia, and are regularly disposed between the radial bars of acanthin, usually so that they are as far removed from them as possible, i.e., in the centre between each three or four bars ; these latter may indeed be regarded as strongly developed axial threads, which have become changed into acanthin (§ 95, A). The soft myxopodia, or pseudopodia without axial threads, are much more numerous than the others, and arise from the sarcodictyum or exoplasmic network which ramifies over the surface of the calymma. Their number and arrangement seem, however, in many (if not in all) ACANTHARIA to be regular and not to possess the extraordinary variability seen in the other three legions. In many Acanthometra the sarcodictyum exhibits a symmetrical con formation, with regular or subregular, polygonal (mostly hexagonal) meshes, and generally the stronger threads of the sarcodictyum secrete a firm, homogeneous or fibrillar, striated substance, which forms a network of ridges on the surface of the calymma. In the Acanthophracta the place of this is taken by the acanthin network of the primary lattice-shell. The axopodia of the Acanthometra are usually about as long as the radial spines between which they stand; their stiff axial thread is surrounded by a soft sheath of protoplasm, communicating with the thin sarcomatrix which surrounds the central capsule. Numerous branches pass into the calymma from the exoplasmic sheath of the axial threads, and form by their interweaving a loose sarcoplegma. The most peculiar differentiated products of the exoplasm of the ACANTHARIA, however, are the myophane fibrillae of the Acanthometra, which have already been described under the name of myophriscs (§ 96).
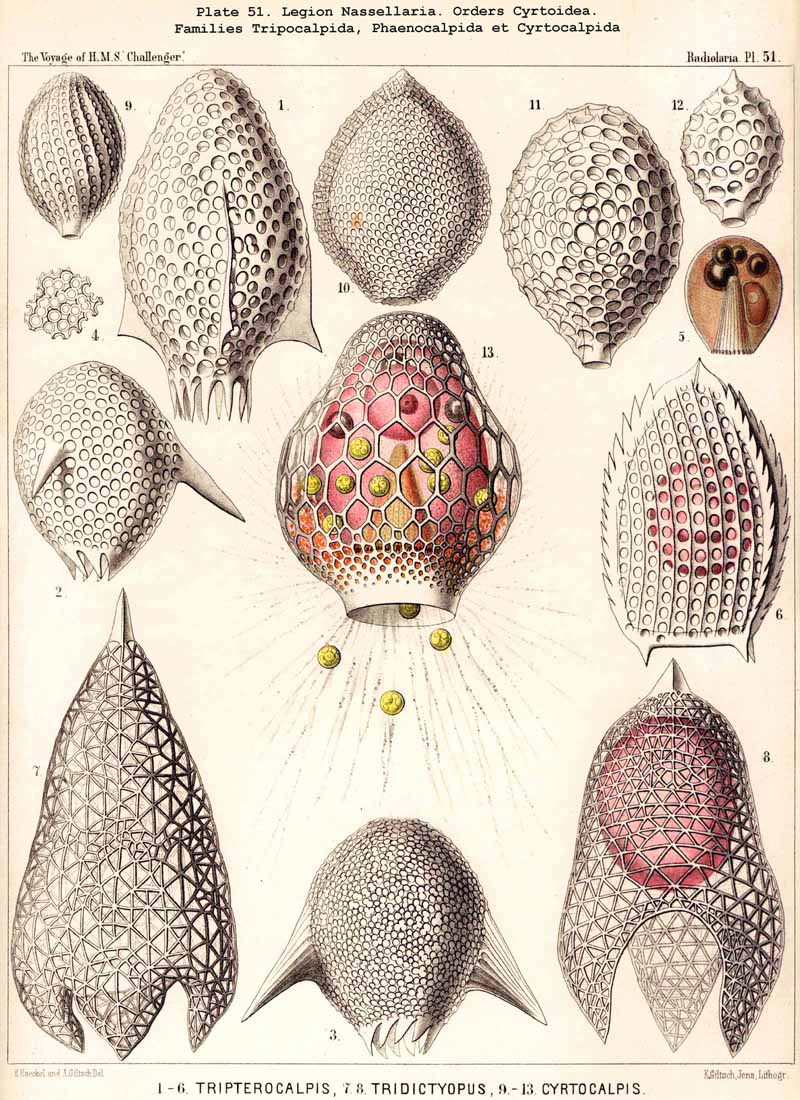
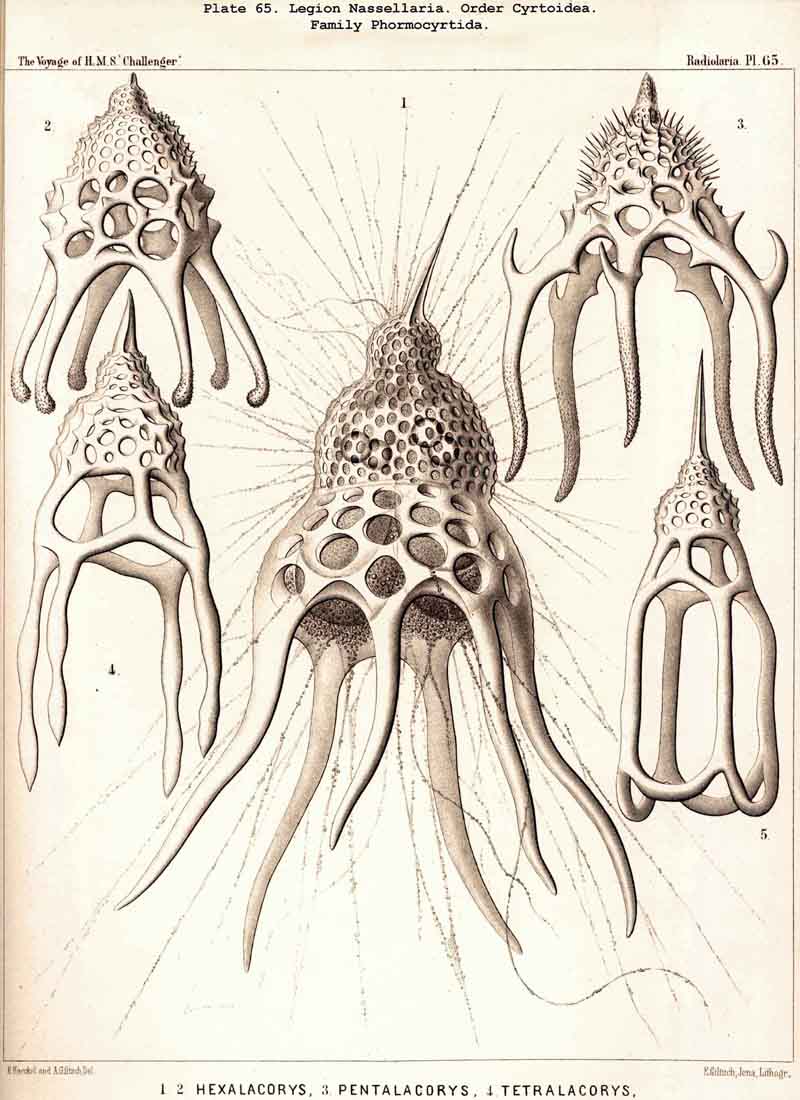
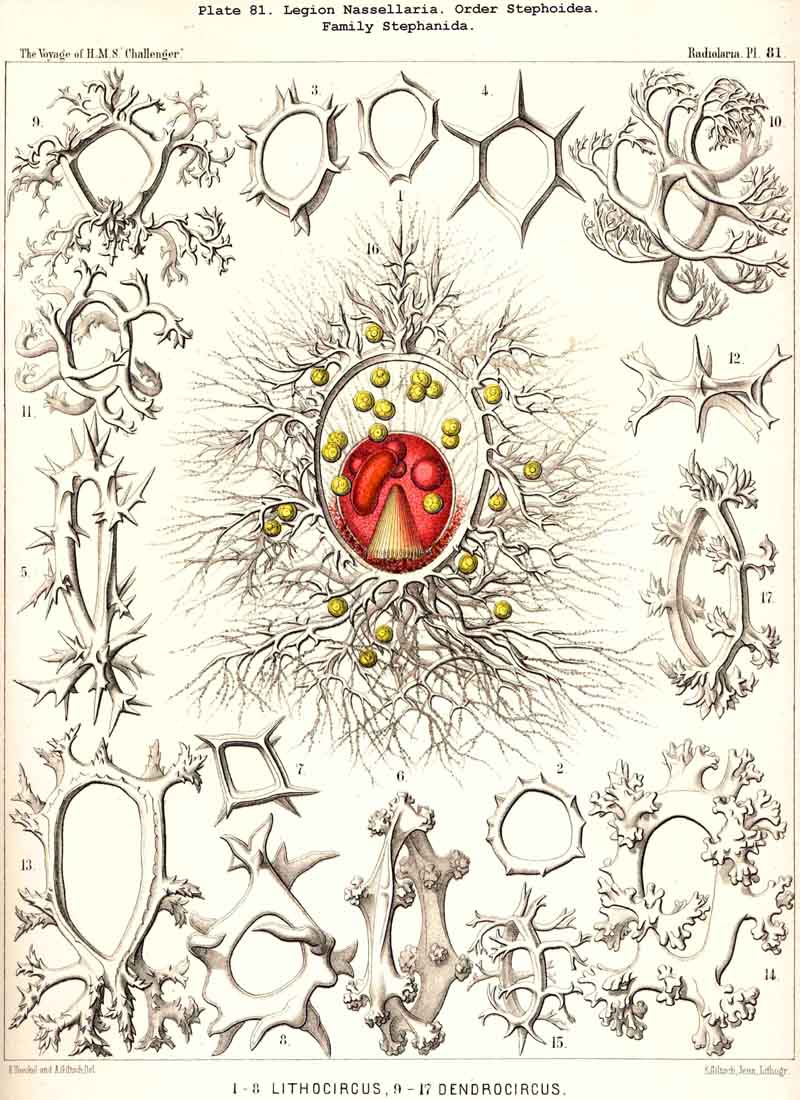
99. The Exoplasm of the Monopylea.-The extracapsular protoplasm of the NASSELLARIA or MONOPYLEA arises only from the porochora, or the intracapsular podoconus, the oral base of which is formed by this porous area. The pseudopodia or protoplasmic threads which pass through the pores of the latter, united into a bundle, are not very numerous (in most NASSELLARIA probably between thirty and ninety), and unite just outside it to form a thick discoid sarcomatrix; this covers the porochora completely below, and spreads out in the form of a thin envelope of exoplasm over the whole surface of the central capsule; at the apical portion of the latter the sarcomatrix is often so thin that it can only be recognised by the aid of reagents; it separates the membrane of the central capsule from the surrounding calymma. The pseudopodia, which penetrate the latter and by loose anastomoses form a wide-meshed sarcoplegma within it, are usually not very numerous. The greater part of them radiate in a bunch downwards from the basal disc of the sarcomatrix, and a smaller number arise from the thinner envelope which covers the remainder of the central capsule (Pl. 51, fig. 13; Pl. 65, fig. 1; Pl. 81, fig. 16). On the outer surface of the calymma the collopodia, which have passed through it, unite to form the sarcodictyum, and through the siicification of this the primary lattice-shell arises in the great majority of the NASSELLARIA. From the surface of the sarcodictyum arise the astropodia, or free pseudopodia which radiate outwards into the water. Their number in most MONOPYLEA is relatively small, but their length appears to be very great.
100. The Exoplasm of the Cannopylea.-The extracapsular
protoplasm of the PHAEODARIA or CANNOPYLEA is much better developed as
regards volume than in the other three legions, and is connected with the
intracapsular sarcode by only a few apertures in the capsule-membrane.
In most PHAEODARIA three of these are present, the astropyle or main-opening
at the oral pole of the main axis, and the two lateral parapylae or accessory
openings on either side of the aboral pole (§ 60). In several families
the latter appear to be wanting, whilst in others their number is increased
; these families have not yet, however, been observed during life. The
protoplasm projects both from the oral main-opening and from the two aboral
accessory openings in the form of a thick cylindrical rod ; the tube into
which each opening is produced in many PHAEODARIA (longer in the case of
the astropyle, shorter in the parapylae) being regarded as an excretion
from this protoplasmic cylinder. The sarcode threads within the tube appear
like a bundle of fibnils, either quite hyaline or finely striated. After
issuing from the mouth of the aperture they pass over into a thick sarcomatrix,
which surrounds the central capsule entirely and separates it from the
enclosing calymma. In the neighbourhood of the basal astropyle the sarcomatrix
is usually swollen into a thick lenticular disc, which is in direct contact
with the peculiar phaeodium of this legion (§ 89). The pseudopodia,
which radiate from the sarcomatrix, and form by anastomosis a wide-meshed
sarcoplegma within the calymma, are usually not very numerous in the PHAEODARIA,
but are very strong. Sometimes two stronger bundles of collopodia may be
distinguished at the two poles of the main axis, an oral bundle (in the
direction of the proboscis of the astropyle) and an aboral bundle (at the
opposite pole between the parapylae). The collopodia of the sarcoplegma
unite at the surface of the calymma into a regular or irregular sarcodictyum,
which, in most PHAEODARIA produces by the secretion of a peculiar silicate
the primary lattice-shell. The free astropodia, which pass outwards from
the sarcodictyum into the water, are in most PHAEODARIA very numerous (Pl.
101,
fig. 10). Since, however, only a few species of this great legion have
been observed in a living state, their pseudopodia require further accurate
examination.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
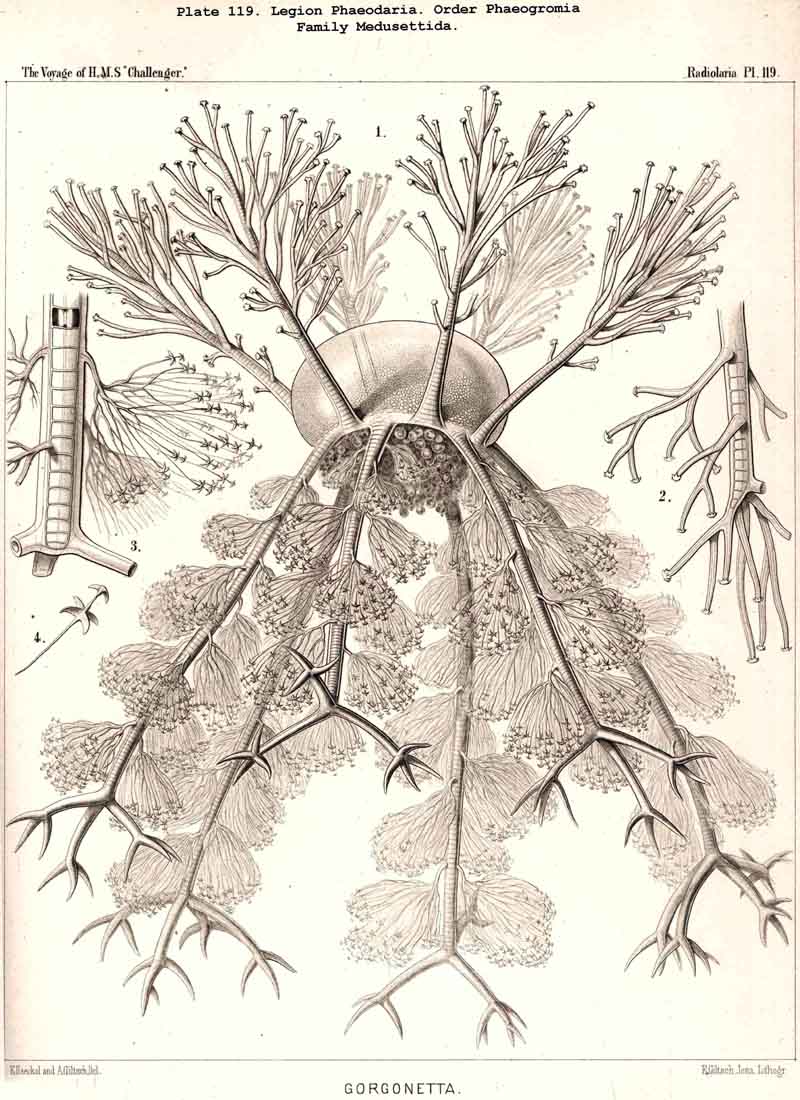{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}