51. Components of the Central Capsule.-In all Radiolaria without exception, at some period of life or other, the central portion of the soft body is separated from the peripheral portion by an independent, anatomically recognisable membrane; this membrane with all its contents is designated the central capsule, and is the peculiar central organ of the unicellular body, which distinguishes the Radiolaria most clearly from the other Rhizopoda. In the great majority of the Radiolaria the volume of the central capsule is less than that of the surrounding peripheral soft body which we place in opposition to it as "extracapsulum." The "capsule-membrane," which separates these two constituents, arises very early in most Radiolaria, and persists throughout their whole life. In some species, however, the membrane only appears later, immediately before the formation of the spores, and hence is absent for a considerable period. Regarded as a whole, then, the capsule consists of the following parts :- (1) the capsule-membrane; (2) the enclosed endoplasm, or intracapsular protoplasm; (3) the nucleus. But in addition, many other non-essential structures may be enclosed in the central capsule, especially hyaline spheres (vacuoles), fatty spheres, pigment granules, crystals, &c.
The central capsule was first described in my Monograph in 1862 (pp. 69-82) as the most characteristic component of the Radiolarian organism, and distinguished from the whole extracapsular soft body. The fact that it has recently been reported as absent by various authors is due to their having observed young or unripe specimens, before the formation of the spores. In some species of Polycyttaria and ACANTHARIA the membrane persists only a very short time.
52. The Primary Form of the Central Capsule.-The form of the central capsule is originally a geometrical sphere; and if in accordance with our monophyletic hypothesis all Radiolaria are to be derived from one common stem-form (Actissa, see p. 12), then the central capsule of this common stem-form must be regarded as perfectly spherical (Procyttarium, p. 13, Pl. 1, fig. 1). Since, further, the enclosed nucleus and the surrounding calymma of this primitive archaic form must also be spheres, and since the nucleus lies in the centre of the body, and the protoplasm is evenly distributed between it and the membrane, it follows that no axes or excentrically differentiated parts are to be distinguished in this most primitive Radiolarian. Rather in the primary central capsule all parts are concentrically and evenly arranged round its centre. This primary spherical form becomes modified in most Radiolaria into various secondary ground-forms, which are correlated partly with the structure of the capsule itself, and partly also with the development of openings in its membrane. In general the ground-form of the central capsule is polyaxon in the Porulosa (SPUMELLARIA and ACANTHARIA); but in the Osculosa centraxon forms are more frequently observed; in the NASSELLARIA the ovoid (allopolar monaxon) form is predominant, and in the PHAEODARIA the rhomboid or amphithect pyramid. In these latter, the astropyle indicates the basal pole of the vertical main axis, whilst the two parapyIae (right and left) mark the poles of the frontal transverse axis. In the NASSELLARIA the centre of the porochora corresponds with the basal pole of the main axis, whilst no transverse axes are originally present.
53. The Secondary Forms of the Central Capsule.-The original purely spherical form of the central capsule persists only in the minority of the Radiolaria, namely, the greater part of the SPUMELLARIA and ACANTHARIA; it passes over into various other secondary forms in the majority of the class, in the whole of the NASSELLARIA and PHAEODARIA, and in a considerable portion of the SPUMELLARIA and ACANTHARIA. These secondary or derived forms may be divided into two quite distinct groups, which may be designated endometamorphic and exometamorphic; in the former the cause of the divergence of the secondary form from the sphere lies in the internal structure of the central capsule; in the latter it lies in the external influence exerted by the growth of the skeleton. Obviously the former series of modifications is more significant than the latter.
54. The Endometamorphic Forms of the Central Capsule.-The secondary forms of the central capsule, which are due to internal causes connected with its growth, are as follows :-
The capsule-membrane was first indicated as the most important and absolutely constant component of all Radiolaria, and as the differential character of the class, in my Monograph (1862, pp. 69-71). The careful investigations of R. Hertwig have confirmed this view and at the same time have yielded the most important conclusions regarding the nature and systematic significance of the openings in the Capsule (op. cit., 1879, pp. 105-107). On the contrary, Karl Brandt has recently propounded the theory that the capsule-membrane is by no means a constant part of the Radiolarian organism, but is lacking in certain species of Collozoum and Sphaerozoum (1881, p. 392). This contradiction is explained by the fact that in some Collodaria and Acanthometra the formation of the central capsule takes place much later than in the other Radiolaria, in some species indeed only just prior to the development of the swarm spores. I have recognised the presence of it in all species which I have investigated (more than a thousand), and even in those in which Brandt denies its existence. It is often very delicate and may easily be overlooked, especially when the contents of the capsule are colourless, but in all cases by the prudent application of staining fluids and other reagents its presence may be demonstrated. Even in those cases in which the contour of the capsule was not visible, and its contents appeared to pass without definite boundary into the matrix of the extracapsulum, it was possible by the use of appropriate stains or reagents, which would not penetrate the capsule, or of those solvents which were capable of dissolving its contents and of causing it to swell up like a distended bladder, to recognise the existence of the membrane. Those Radiolaria in which it is truly absent are young animals of species in which the membrane is only formed immediately before sporification, and persists but for a short time (e.g:, species of Collozoum, Sphaerozoum, Acanthometra, Acanthochiasma, &c.).
57. The Capsule-Openings of the Peripylea (or Spumellaria).The capsule-membrane of the PERIPYLEA is generally perforated by extremely fine and numerous pores, which are distributed at equal distances over the whole surface, and are precisely alike in all parts of the capsule. Hence the SPUMELLARIA may be called "Holotrypasta" or "Porulosa"; they agree with the ACTIPYLEA in being devoid of an osculum or operculum; they are distinguished from the latter group mainly in that their pores are equally distributed over the whole surface of the capsule, whilst in the ACTIPYLEA the pores are disposed in definite groups or lines, separated by large imporous areas.
The central capsule of the SPUMELLARIA, with its innumerable fine and evenly distributed pores, must be regarded as the primitive arrangement, from which the different central capsules of the three other legions have been developed. The central capsule of the ACTIPYLEA has been derived from that of the PERIPYLEA by reduction in the number of the pores and their distribution in definite, regularly disposed areas in the membrane. The Central capsule of the Osculosa is characterised by the formation of a special main-aperture (osculum) at the basal pole, which is closed in the MONOPYLEA by the porochora, and in the CANNOPYLEA by the astropyle; the remaining pores, with the exception of the accessory openings of many CANNOPYLEA, remain undeveloped in both these legions. In the same way Hertwig regards the central capsule of the PERIPYLEA as the primitive form (1879, L. N. 33, p. 107).
58. The Capsule-Openings of the Actipylea (or Acantharia).-The capsule-membrane of the ACTIPYLEA is perforated by very numerous fine pores, which are regularly distributed over the surface of the central capsule, and separated by imporous intervals. Hence the ACANTHARIA belong to the "Holotrypasta" or "Porulosa"; they have neither osculum nor operculum, and agree in this particular with the PERIPYLEA; but they are separated from these latter chiefly by the fact that their pores are much less numerous, and marked off into regularly arranged groups or lines by imporous intervals. In the PERIPYLEA, on the contrary, the pores are much more numerous and are evenly distributed over the whole surface of the capsule.
The central capsule of the ACANTHARIA has hitherto been for the most part confounded with that of the SPUMELLARIA, and no clear distinction has been drawn in this respect between the two legions of the Porulosa. Hertwig, who in 1879 first discovered the remarkably different structure of the Osculosa (NASSELLARIA and PHAEODARIA), recognised no distinction between the structure of the capsules in the PERIPYLEA and ACTIPYLEA (his Acanthometrea), and supposed that in both these legions "very fine pores were evenly distributed in large numbers over the capsule-membrane" (loc. cit., p. 106). I have, however, during the last few years convinced myself, by the careful comparative investigation of numerous ACANTHARIA, that in this respect they are quite distinct from the SPUMELLARIA (with perhaps the exception of the Astrolophida, which are nearly related to the primitive Actissa). The number of pores in the ACTIPYLEA is usually very much smaller than in the PERIPYLEA, and they are regularly arranged in groups.
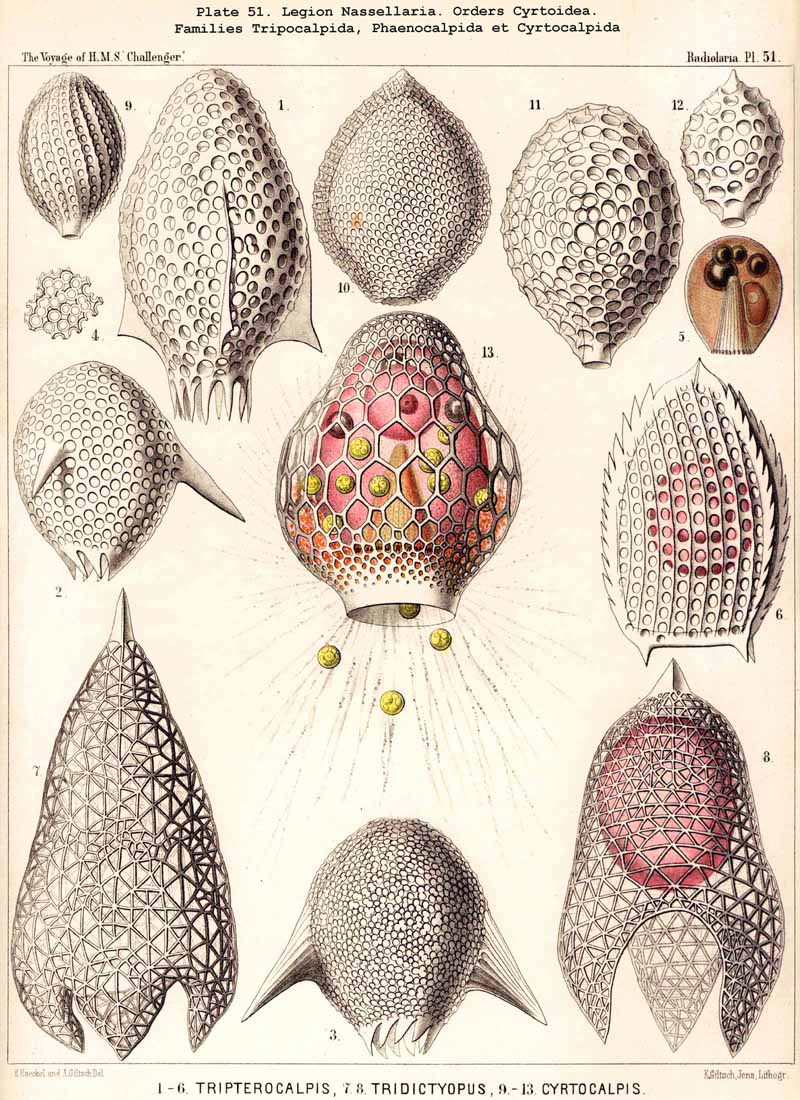
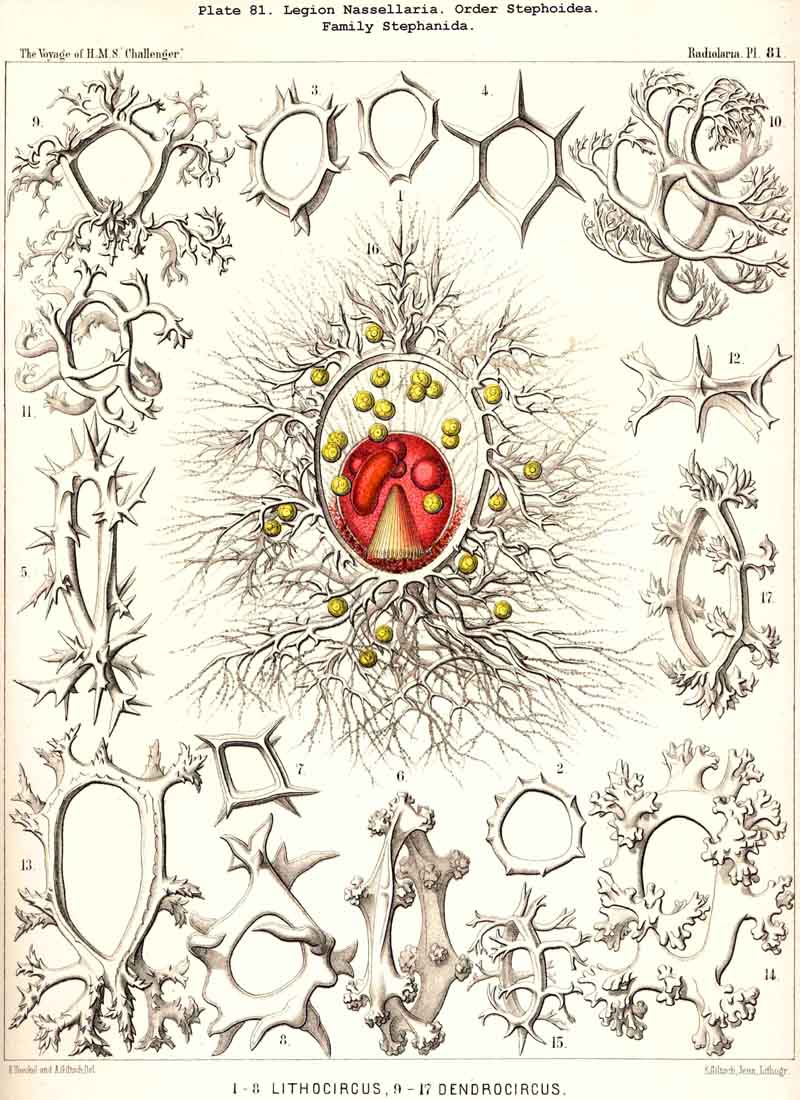
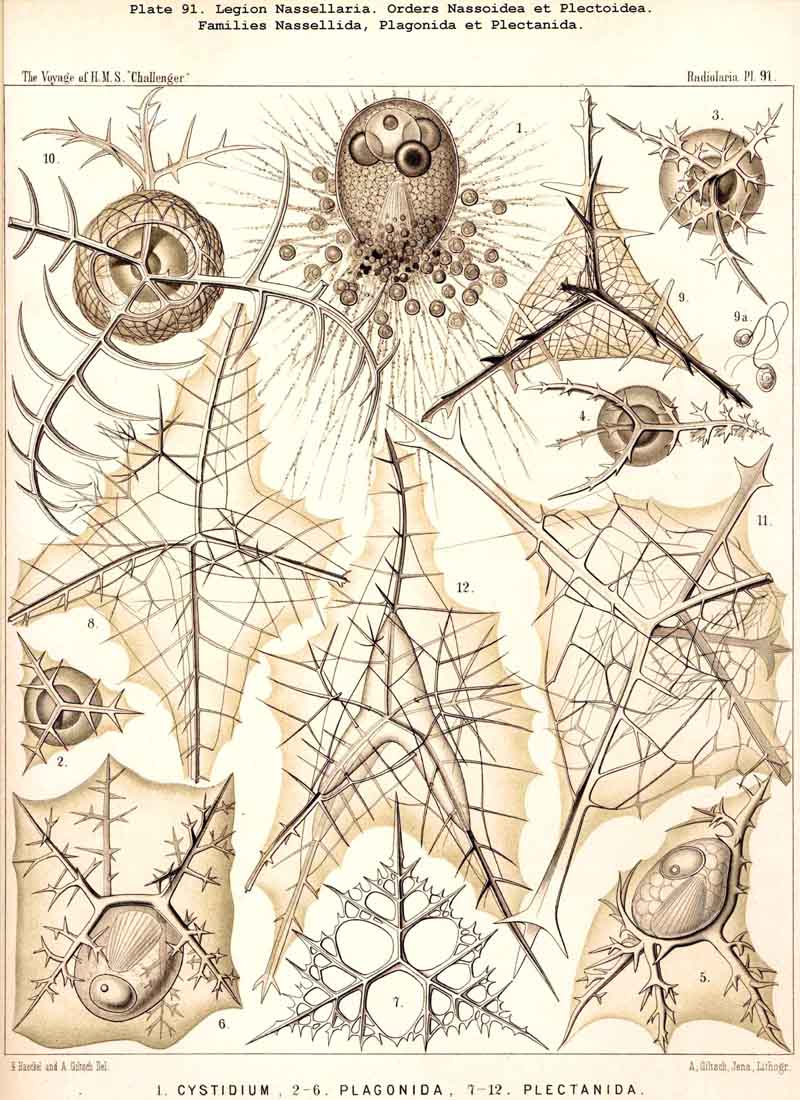
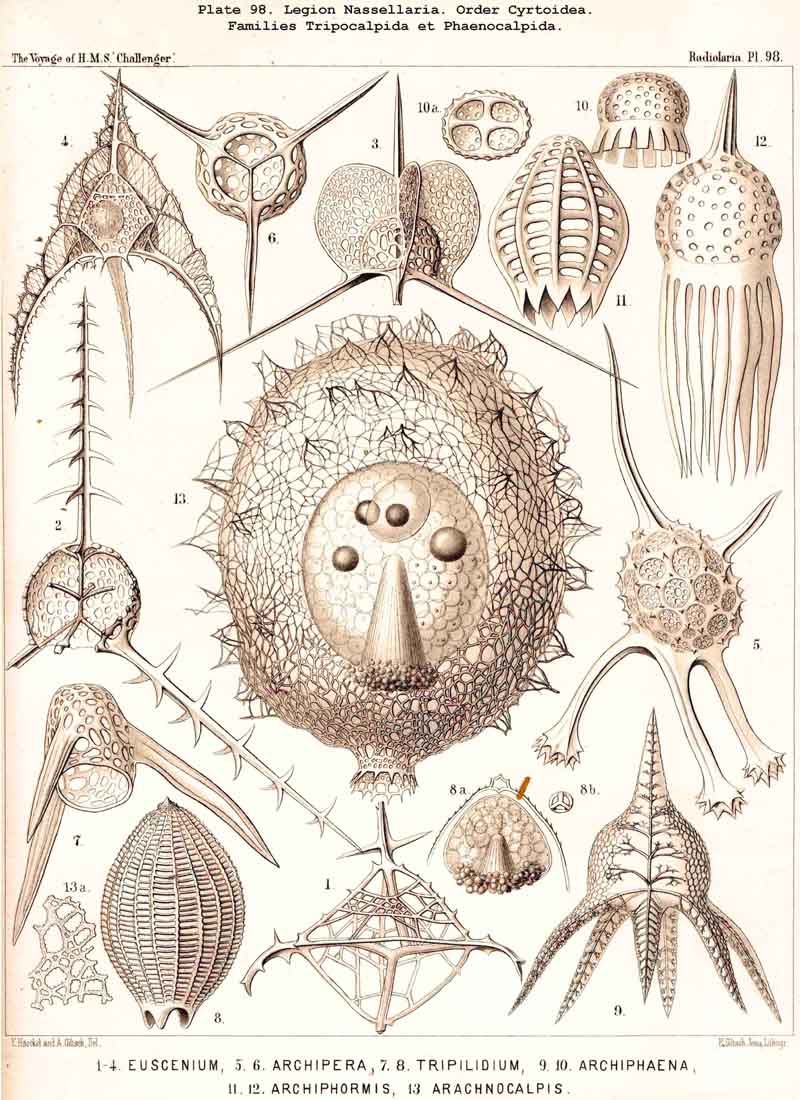
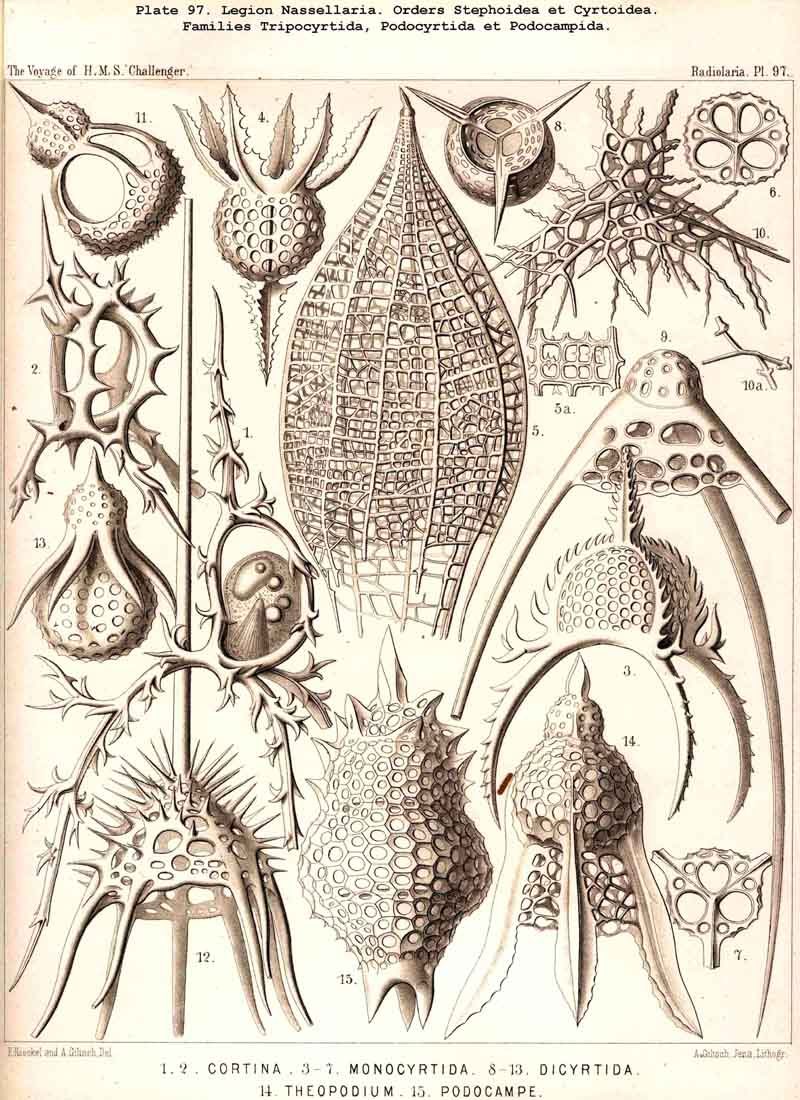
59. The Capsule- Openings of the Monopylea (or Nassellaria.)The capsule-membrane of the MONOPYLEA always possesses a single large main-opening, an osculum, which lies at the basal pole of the main axis, and is closed by a circular perforated lid (operculum porosum). When seen from the surface this lid appears as a clearly defined porous area (porochora or area porosa), and forms the horizontal base of a peculiar cone, which stands vertically in the interior of the capsule and may be designated the " thread-cone" (podoconus). The NASSELLARIA may hence be termed "Merotrypasta" or "Osculosa," like the CANNOPYLEA; the structure and significance of the circular lid (operculum), which closes the main-opening (osculum) is, however, quite different in the two legions. Whilst the lid of the CANNOPYLEA (astropyle) is solid, traversed by radial ribs, and only perforated in its centre by a short tube (proboscis), in the MONOPYLEA the operculum (porochora) is always perforated by numerous vertical fine pores, and is in connection with the peculiar internal "pseudopodial cone" (podoconus, Pl. 51, figs. 5, 13; Pl. 81, fig. 16; Pl. 91, fig. 5; Pl. 98, fig. 13). The pores are separated by small vertical, highly refractive rods (opercular rhabdillae); these become intensely stained by carmine, and are either evenly distributed over the surface of the porochora or arranged in definite groups. The outer or distal end of each rod is rounded, sometimes thickened like a club or split into lobes; the inner or proximal end is usually pointed, and stands in connection with a myophane thread of the podoconus (see § 79). The primary circular form of the porochora, in which the opercular rhabdillae are evenly distributed in a horizontal plane, undergoes various secondary modifications in many NASSELLARIA. The triradial structure of the skeleton, which characterises the majority of the legion, causes a splitting of the base of the central capsule into three or four lobes; this division also affects the porochora, which lies in the centre of the base, so that the rhabdillae become arranged in three or four equal circles. If, however, the lobes of the central capsule become larger and protrude through the three or four collar pores of the cortinar septum, the central porochora may separate entirely into three or four elongated tracts, which lie on the axial side of the magnified lobes; the rhabdillae are then arranged over the whole surface of these tracts, on the outer aspect of which run the longitudinal myophane fibrillae of the podoconus (compare §§ 79 and 99).
The porous area of the MONOPYLEA was first described by Hertwig in 1879,
and shown to be the characteristic main-opening of the central capsule
in various families belonging to this legion (L. N. 33, pp. 71, 73, 83,
106, Taf. vii., viii.). According to his view "the capsule-membrane in
the porous area becomes thickened around each pore into a rod, perforated
by a canal," and the intracapsular protoplasm passes outwards through these
fine canals (loc. cit., p. 106). I am not able to share this interpretation,
but think rather that I have convinced myself by the examination of some
living NASSELLARIA, and of many well-stained and preserved preparations
in the Challenger collection, that the rods are solid, specially
modified portions of the capsular wall, and that the protoplasm does not
pass through them but through pores which lie between them.
60. The Capsule-Openings of theCannopylea (or Phaeodaria).- Thecapsule-membrane of the CANNOPYLEA always possesses only a single large main-opening or osculum, which lies at the basal pole of the vertical main axis, and is closed by a circular radiated lid (operculum radiatum). This operculum appears, when seen from the surface, as a sharply defined stellate area (astropyle), from the middle of which arises a shorter or longer cylindrical tube, the proboscis. Hence the PHAEODARIA, like the MONOPYLEA, belong to the "Merotrypasta" or "Osculosa"; the structure and significance of the circular operculum, which closes the main-opening (osculum), are, however, quite different in the two legions. Whilst the operculum of the MONOPYLEA (porochora) is perforated by numerous fine vertical pores, and connected with the peculiar internal pseudopodial cone (podoconus), this structure is entirely wanting in the CANNOPYLEA, and instead of it there is a solid operculum, with radial ribs which originate at the base of its central tubular mouth; this tube (proboscis) is cylindrical, often conical at the base, of very variable length and with a round aperture at either end. In spite of the great difference which the various families of CANNOPYLEA exhibit in the formation of their skeleton and its appendages, the constitution of this characteristic stellate main-opening (astropyle) is always essentially the same; both the stellate operculum itself, and the proboscis which rises from its centre, show only slight differences in the various groups. In addition to this large main-opening most PHAEODARIA possess several small accessory openings (parapylae); and usually two of these are present, placed symmetrically right and left of the aboral pole of the main axis and in the frontal plane (Pl. 101, figs. 2, 6, 10; Pl. 104, figs. 1, 2a). Sometimes there are more numerous accessory openings (three to six or more) regularly arranged, as in the two peculiar families, Circoporida and Tuscarorida; occasionally also there is only a single parapyle, at the aborab pole of the main axis (e.g., in Tuscaridium). The parapylae seem to be quite absent in the families Challengerida, Medusettida, Castanellida, and perhaps also in other PHAEODARIA. The form and structure of the small accessory openings appear to be always the same. The outer capsule-membrane is elevated in the form of a short cylindrical tube or "apertural ring" (collare paraboscidis), the external margin of which bends inwards, and at the base of the ring passes over into the delicate internal capsule membrane. Upon this apertural ring is situated a longer or shorter "apertural cone" (paraboscis), which is a tubular, cylindrical or conical, prolongation of the membrane, open externally.
The peculiar capsule-openings of the PHAEODARIA were first discovered and carefully described by Hertwig in 1879 (L. N. 33, pp. 95, 107). He found in all the six genera which he examined three openings, a main-opening at the basal pole of the main axis and two accessory openings, one on either side of the apical pole; hence he named the whole group "TRIPYLEA." This name, however, is not applicable to the numerous PHAEODARIA mentioned above, which have only a main opening without any accessory openings, nor to those genera in which the number of the latter is variable. I have, therefore, replaced Hertwig's designation by the term "CANNOPYLEA," which has reference to the peculiar tubular form of the opening. This I find much more developed in many PHAEODARIA than Hertwig has represented, and I must also, in certain particulars, dissent from his delineation of the minute structure, although this is in the main remarkably accurate.
61. The Nucleus.-The nucleus, enclosed in the central capsule of all Radiolaria, behaves in every respect like a true cell-nucleus, and thus lies at the base of the now universal opinion, that the whole Radiolarian organism, in spite of its varied development and remarkable variations, is unicellular and remains throughout life a true individual cell. This important theory is not invalidated by the fact that the nucleus undergoes peculiar modifications in many groups, and in certain groups presents appearances seldom or never seen elsewhere.
62. Uninuclear and Multinuclear Radiolaria (Monocaryotic and Polycaryotic).- All Radiolaria present two different conditions in respect of the behaviour of the nucleus, since in their young stages they are uninuclear (monocaryotic), and in later stages multi-nuclear (polycaryotic). This is readily explained by the fact that each individual Radiolarian is developed from a simple unicellular swarm-spore, and that afterwards, before the formation of swarm-spores, the single nucleus divides into many small nuclei. Thus in the Radiolaria the nucleus is pre-eminently the organ of reproduction and inheritance. The division of the originally single nucleus into many small nuclei may take place, however, at very different periods, so that the Radiolaria may be divided in this respect into precocious and serotinous.
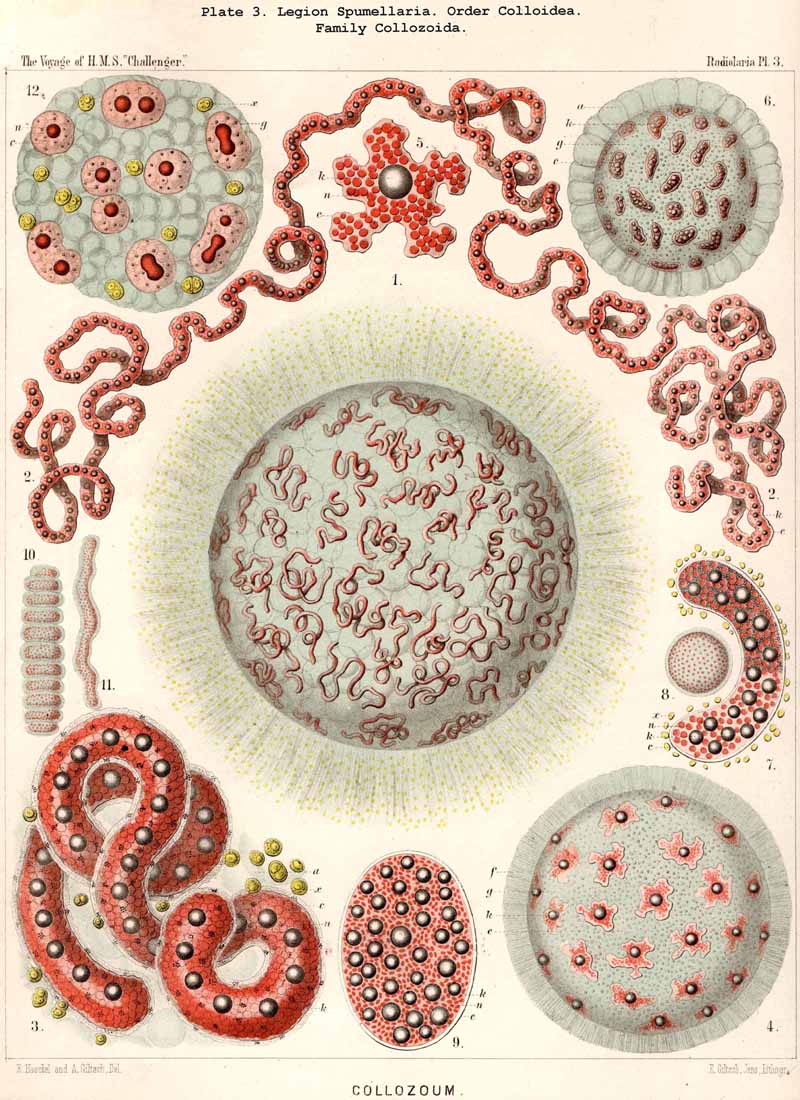
63. Serotinous and Precocious Radiolaria.In the great majority of the Radiolaria the division of the nucleus takes place only at a late period, a short time or even immediately before the process of spore formation; it then breaks up rapidly into numerous small nuclei (always more than one hundred, sometimes many thousands), and each of these either becomes itself the nucleus of a swarm-spore, or by repeated division gives rise to a group of spore-nuclei. All those Radiolaria which are uninuclear during the greater part of their existence, and in which the process of division is late, and takes place rapidly, are called "serotinous "or late-dividing forms. To this category belong all PHAEODARIA and NASSELLARIA, as well as all the solitary or monozoic SPUMELLARIA and some ACANTHARIA. On the other hand, the name "precocious," or early dividing, is applied to those Radiolaria in which the division of the nucleus takes place very early, and in which, therefore, the cell is multinuclear during the greater part of its existence. This is the case in all the social or polyzootic Radiolaria (Polycyttaria, Pls. 3-8), and also in the great majority of the ACANTHARIA, both Acanthometra and Acanthophracta. In the last two groups, however, there are numerous exceptions, and these are seen in remarkably large species, characterised by the great size of the central capsule. From a phylogenetic point of view, the conclusion is allowable that the precocious forms are secondary, and have arisen by adaptive modification from the primitive serotinous stem. In the Polycyttania (or social SPUMELLARIA, i.e., the three families Collozoida, Sphaerozoida, and Collosphaerida), the cause of the adaptation lies most probably in the formation of the colony itself, for all these three families are so closely related to three corresponding families of serotinous, monozootic Radiolania (Thalassicollida, Thallassosphaerida, Ethmosphaerida), that certain species of the latter are hardly to be distinguished from isolated individuals of the former. Perhaps the remarkable formation of the large central oil globule, which particularly characterises the Polycyttaria, is the prime cause of their early nuclear division. In the ACANTHARIA the cause is most likely to be found in the characteristic centrogenous development of their acanthin skeleton, whose radial bars first of all appear in the centre of the capsule. Hence arises directly the excentric position of the nucleus, which in the archaic stem of ACANTHARIA (Actissa?) was probably central. In any case, but little weight is to be laid upon the precocious division of the nucleus in the ACANTHARIA in general, inasmuch as in certain species (both Acanthometra and Acanthrophracta) the more usual serotinous division persists.
64. Central and Excentric Nuclei.-The position of the nucleus in the interior of the central capsule was no doubt primitively central, and this situation in the geometrical centre of the original spherical central capsule has been accurately retained in all monozootic SPUMELLARIA; in the polyzootic families of this legion (Polycyttania), on the contrary, it is obscured by the precocious division of the nucleus. In the other three legions, which may be phylogenetically derived from the SPUMELLARIA, the position of the nucleus is rarely central, but usually excentric, or at most subcentral. In the ACANTHARIA. (both Acanthometra and Acanthophracta) the central position of the nucleus is at once excluded by the constantly centrogenous development of the skeleton; the nucleus is therefore always excentric, and may lie at either side; it usually divides very early into numerous separate nuclei, which are usually distributed in the peripheral portions of the central capsule. In the NASSELLARIA the development of the porochora, and of the podoconus which stands upon it, brings about the formation of a vertical axis, and in consequence the central capsule assumes a monaxon form (usually ovoid or conical); the nucleus then lies in the main axis, but excentrically between the apex of the podoconus and the aboral pole. In many NASSELLARIA, however, especially when the podoconus is so large that its apex approaches the aboral pole of the central capsule, the nucleus is pressed to one side and lies quite excentrically. The PHAEODARIA exhibit a different arrangement; the large spheroidal nucleus is always subcentral, so that its main axis corresponds with that of the concentric spheroidal central capsule; but since the astropyle always occupies the oral pole of the latter, and since the distance of the nucleus from this pole is always somewhat different from its distance from the other, it follows that, strictly speaking, the nucleus never lies accurately in the geometrical centre.
65. Homogeneous and Allogeneous Nuclei.-The nucleus of the Radiolaria not only exhibits a similar structure and composition, and suffers similar modifications to those which are found to occur in the case of other cell-nuclei, but also to some extent shows very peculiar developmental forms, which are seldom or never found in other cells. In the first place the nuclei may be divided into homogeneous and allogeneous, the former are structureless and consist of a uniform mass of nuclein, whilst the latter are composed of different substances and show various structural relations. Homogeneous nuclei, whose whole mass is uniform and exhibits no structural differentiation, are probably always to be found in the swarm-spores; in the fully developed Radiolarian body they are found only in the first legion, SPUMELLARIA, and that both in many Monozoa (especially small Sphaeroidea and Prunoidea) and in the Polyzoa (or Polycyttaria). The whole mass of these homogeneous nuclei, which are usually spherical or ellipsoidal, consists of uniform, perfectly clear and transparent nuclein, and becomes evenly stained by carmine, haematoxyline, &c. They may be readily distinguished by these means from the clear vacuoles or "hyaline vesicles," which are evenly distributed in the endoplasm of many Radiolaria, and may be confused with the former. Allogeneous nuclei, which are always composed of different parts and often show complicated structural relations, are found developed in the great majority of Radiolaria. The most important differentiation exhibited by these secondary forms is the separation of the nuclear mass into a firm nuclear substance (caryoplasm) and a fluid nuclear juice (caryolymph). In addition in each nucleus a nucleolus is visible, and often several or many may be seen (see §§ 67 to 70).
66. The Form of the Nucleus.-The nucleus of the Radiolaria shows greater variations in form and structure than are to be found in the majority of cell-nuclei; exception must, however, be made in the case of many animal ovicells, which, in their peculiar form and composition, often recall large Radiolarian nuclei. With respect to the external shape two main forms may be distinguished, as primary and secondary. The primary form of the Radiolarian nucleus is the sphere; it occurs not only in most swarm-spores, but also in most adult forms belonging to the legion SPUMELLARIA, and in individual instances in other groups; indeed the nuclei of most SPUMELLARIA, as also the concentric central capsules in which they lie, are true geometrical spheres. The secondary forms of the nucleus are found in the majority of adult Radiolaria, and arise from the primary spherical forms in various ways, either by the elongation or contraction of one axis, or by the formation of apophyses or processes. The most important of these secondary forms are as follows :-
Both the simple serotinous nucleus of the monozootic SPUMELLARIA, and the numerous precocious nuclei of the Polycyttaria, were first described in my Monograph in 1862, the former as the "endocyst" (" Binnen-Bläschen "),the latter as "spherical transparent vesicles" (" Kugelige wasserhelle Bläschen "). I was in error, however, in regarding the latter as identical with the so-called "hyaline spherules" in the central capsule of many Monozoa, which rather belong to the category of intracapsular vacuoles (see § 72). The credit of recognising, by the aid of the modern methods of staining the distinctness of these two structures, which may readily be mistaken for each other, and of demonstrating the true nature both of the serotinous and precocious nuclei, belongs to Richard Hertwig (1879, L. N. 33).
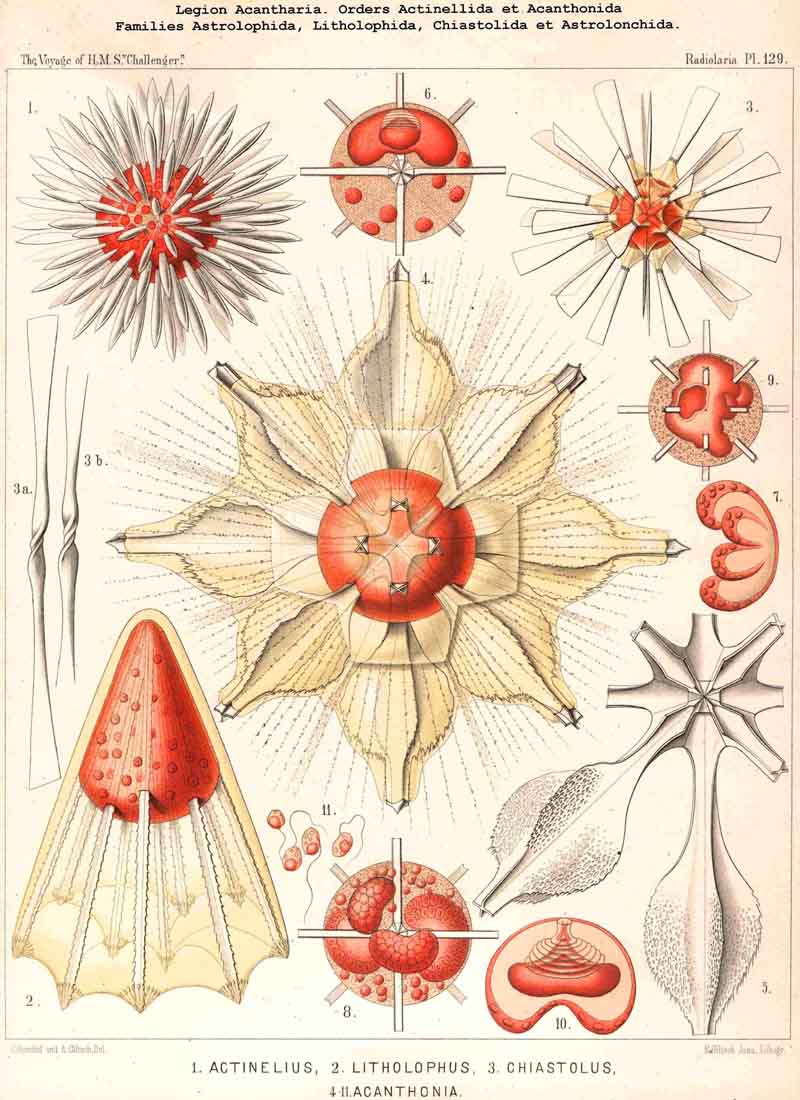
68. The Nucleus of the Actipylea.-The nucleus of the ACANTHARIA or ACTIPYLEA shows very peculiar relations in respect of structure and division, particularly special forms of lobular budding, which belong to the characteristic peculiarities of this singular legion, and are not found among other Radiolaria. The position of the nucleus is always excentric, even in the youngest ACANTHARIA, for the centrogeneous formation of the skeleton, the constant development of the earliest radial portions of it in the middle of the central capsule, forces the nucleus from its normal central position. The majority of the ACANTHARIA, like most Polycyttaria, are precocious, the primary nucleus early dividing into numerous small nuclei (see note A below). Nevertheless there are many exceptions to this rule in different families, e.g., Stauracantha, Xiphacantha, Phatnacantha, and Pristacantha among the Acanthometra, and Stauraspis, Echinaspis, Dodecaspis, and Phatnaspis among the Acanthophracta. In these instances the primary nucleus remains for a long time as a simple excentric ellipsoidal or irregularly round body, even in the fully developed stage, and only at a very late period (sometimes just before the formation of the spores) divides into many small nuclei. Since this serotinous division of the nucleus takes place in different genera of very various groups, it can only be decided by further investigations how widely it is spread among the ACANTHARIA, and upon what circumstances it is dependent (see note B). The division of the nucleus appears to be precocious in the majority of this legion, and a number of small nuclei appear to be early formed by a peculiar process of budding; in most fully developed ACANTHARIA these are disposed in one or two layers under the surface of the central capsule, but if their numbers increase to any considerable extent, the whole space between the skeletal rods becomes filled with small nuclei; sometimes these are homogeneous, sometimes vesicular, 0 .002 to 0.012 mm. in diameter; usually they are spherical and have a small nucleolus (compare Pl. 129, figs. 6-11, and note C).
A. The numerous nuclei, which are to be found in the Central capsule of most mature ACANTHARIA, were first described in my Monograph (1862) as "spherical, transparent vesicles, provided with a small dark granule" (p. 374, Taf. xv. figs. 2, 5; Taf. xvi. figs. 2, 4; Taf. xxi. fig. 7, &c.). Their more minute constitution and peculiar origin were first accurately delineated by R. Hertwig (1879, loc. cit., pp. 11-24, Taf. i-iii.).
B. The fact that in a number of ACANTHARIA the nucleus does not divide early as in the majority of the legion, but only at a later period, was first observed by R. Hertwig in a species of Acanthometra (Xiphacantha serrata), and a species of Acanthophracta (Phatnaspis mülleri = Haliommatidium mülleri) (loc. cit., pp. 11 and 27). This serotinous division of the nucleus seems, however, to be rather widely spread in both sublegions of the ACANTHARIA; I have found, not only in the forms above mentioned, but also in several others belonging to different genera, a single large excentric nucleus, even in those individuals in which the skeleton was fully developed.
C. The peculiar mode of nuclear budding, by which these small nuclei arise, appears to proceed in the following manner (Pl. 129). The vesicular primary nucleus, which, in consequence of the Centrogeneous development of the skeleton protrudes as it grows into irregular lobes (Pl. 129, fig. 9), assumes a peculiar concavo-convex form, sometimes that of a hood or dish, sometimes that of a kidney or sausage. The convex surface is apposed to the capsule-membrane, while the concave is turned towards the central star of the skeleton (fig. 6). There is now formed at the centre of the convex surface of the strong, doubly-contoured, nuclear membrane, a flask-shaped invagination with a narrow neck and expanded base; the membrane now becomes disposed in peculiar folds, which at the narrow aperture of invagination appear as folds, but on the expanded body of the flask take the form of concentric rings, laid closely side by side (Pl. 129, fig. 10). The convex bottom of the flask, which is directed towards the concave proximal side of the nucleus, becomes again invaginated by a central conical apophysis of the enlarged nucleolus, which is situated between them. Usually the nucleolus has already become flattened into a lentiform shape, and upon its distal face a conical apophysis has been developed, which is divisible into a darker proximal and clearer distal portion. The tip of the latter appears to be in direct connection with the nuclear membrane at the centre of the base of the flask-shaped invagination (figs. 6, 10). At this stage of development the nucleus of the ACANTHARIA generally presents the characteristic form of a hood-shaped, concave-convex vesicle, whose radial axis is also the axis of the flask-shaped distal invagination, and of the depressed conical nucleolus, which lies between the latter and the concave side of the nucleus. After this peculiar invagination has persisted for some time in connection with the enlarged nucleolus, both disappear, and then a remarkable growth of lobular processes takes place on the concave proximal side of the hood or kidney-shaped nucleus; from four to eight knobs of unequal size usually appear, and their thickened wall encloses a variable number of small nucleoli; these are at first few but afterwards more numerous (fig. 7). Subsequently these knobs or lobes become completely separated by constriction from the original central mass of the nucleus, and appear as so many separate independent "sausage-shaped bodies" in the hollow central capsule (fig. 8). Each of the bodies now appears, and at first on its convex aspect, to form a large number of small nucleoli, which either separate by constriction from it or become free by its breaking up and lie in numbers in the central capsule. Finally the buds or lobes of the nucleus break up entirely into such nucleoli, which are evenly distributed in the central capsule, and become the nuclei of the swarm- spores (fig. 11). Compare R. Hertwig, L. N. 33, Taf. i.-iii. pp. 19-25.
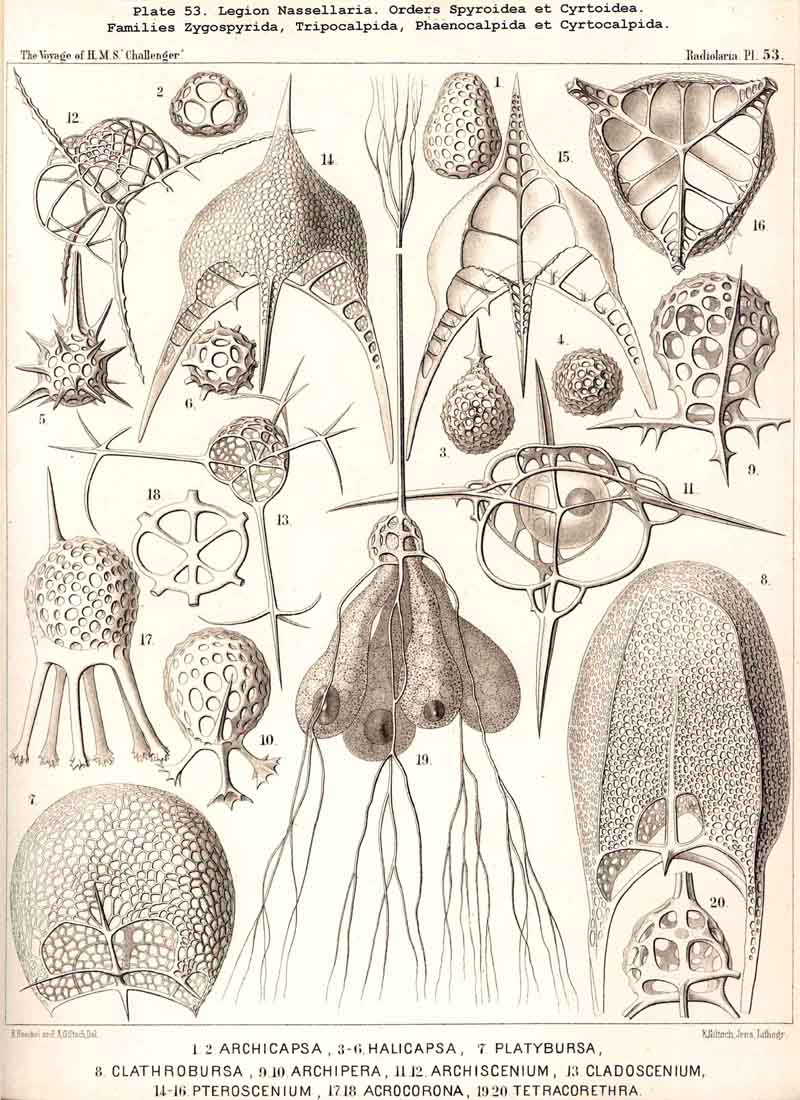
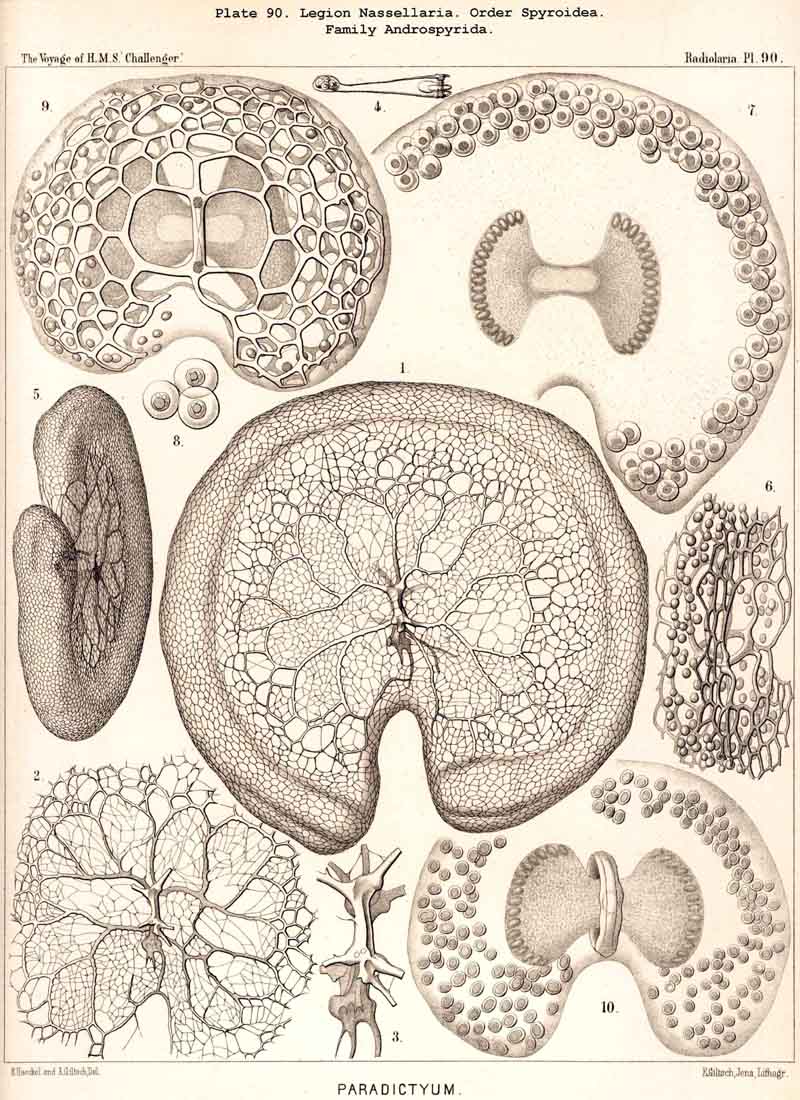
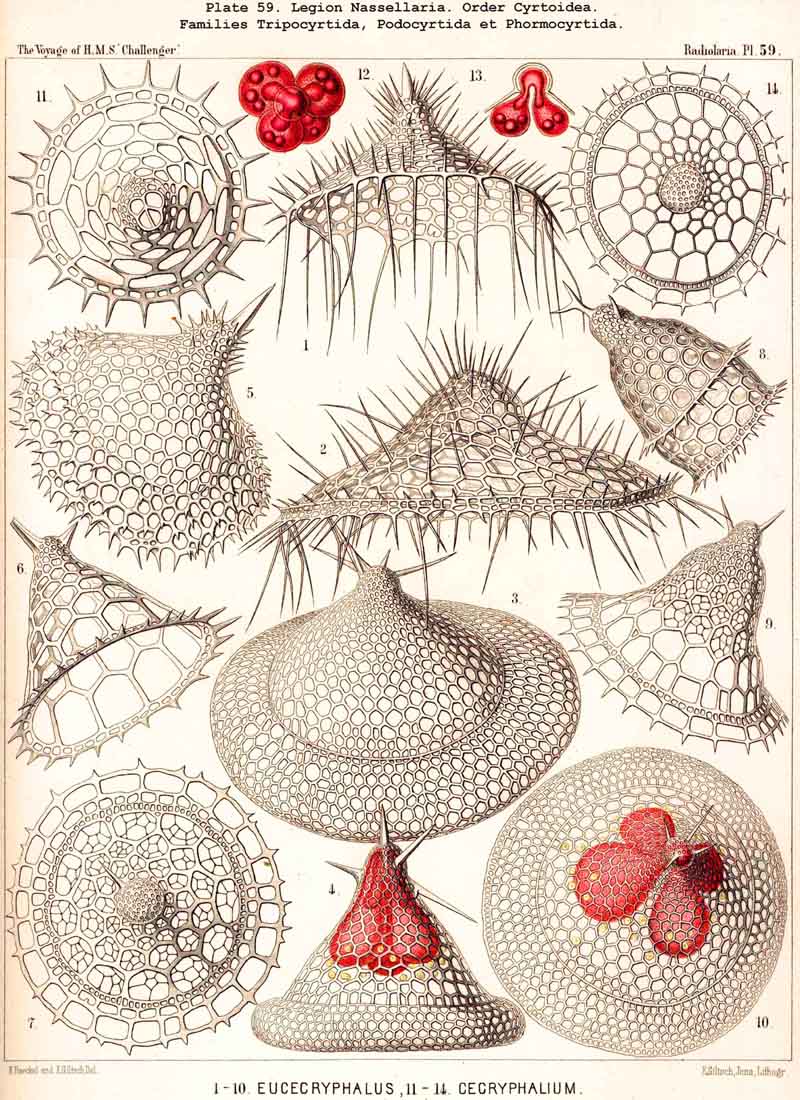
69. The Nucleus of the Monopylea.-The nucleus in the mature forms of the NASSELLARIA or MONOPYLEA is generally simple or lobate, homogeneous or vesicular and excentric, and appears only to divide into numerous small nuclei just before the formation of the spores. Nevertheless I have sometimes, though not often, seen in representatives of very various families of the MONOPYLEA, the central capsule filled with many small spherical homogeneous nuclei (Pl. 53, fig. 19). Hence all the families of this legion appear to be serotinous, their simple primitive nucleus persisting for a long period. It is commonly placed excentrically, and most usually in the apical or aboral portion of the central capsule, either between its apex and the podoconus, or quite excentrically on the dorsal aspect. The simple nucleus of the NASSELLARIA usually appears to be vesicular and to possess a somewhat firm membrane, clear contents, and a rather large, dark coloured nucleolus. In many NASSELLARIA the nucleus is spherical or ellipsoidal (Pl. 53, fig. 11); whilst in many Stephoidea and Spyroidea, where the central capsule is constricted by the sagittal ring and divided into two symmetrical lateral lobes, the nucleus partakes of the same mode of growth, and appears in the middle of the capsule as a transversely placed ellipsoid or even as a short cylinder (Pl. 90, figs. 7, 9), The most remarkable modification in the form of the nucleus is to be found in the multi-articulate Cyrtoidea. Here it is usually enclosed in the cephalis and is spherical, ellipsoidal or spheroidal, often flattened almost into a disc. If now the central capsule increase greatly in size and put forth three or four clavate lobes which hang down through the pores of the cortinar septum into the thorax (or even into the succeeding joints), the nucleus usually undergoes similar modification, and three or four finger-like apophyses are developed from its base, which project into the corresponding lobes of the central capsule (Pl. 59, figs. 4, 12, 13).
The numerous small, spherical, homogeneous nuclei which are to be found in the central capsules of those NASSELLARIA, which are ripe and about to develop spores, were described in 1862 in my Monograph, as "numerous, small, transparent, spherical cells" in the case of various Cyrtoidea (Arachnocorys, Lithomelissa, Eucecryphalus, Eucyrtidium, &c. (loc. cit., pp 302, 305, 309, 321, &c.), and I find them of the same form and dimensions, but deeply stained with carmine in many preparations in the Challenger collection. R. Hertwig has delineated them very accurately in the case of Tridictyopus(1879, loc. cit., p. 84, Taf. vii. fig. 3). He was also the first to recognise the uninucleate condition of the NASSELLARIA, which is much more frequently observed than the serotinous multinucleate condition, and he described very clearly the peculiar lobed nuclei which arise in Cyrtoidea, owing to the protrusion of the nucleus through the cortinar septum (loc. cit., p. 85, Taf. viii. figs. 3-8).
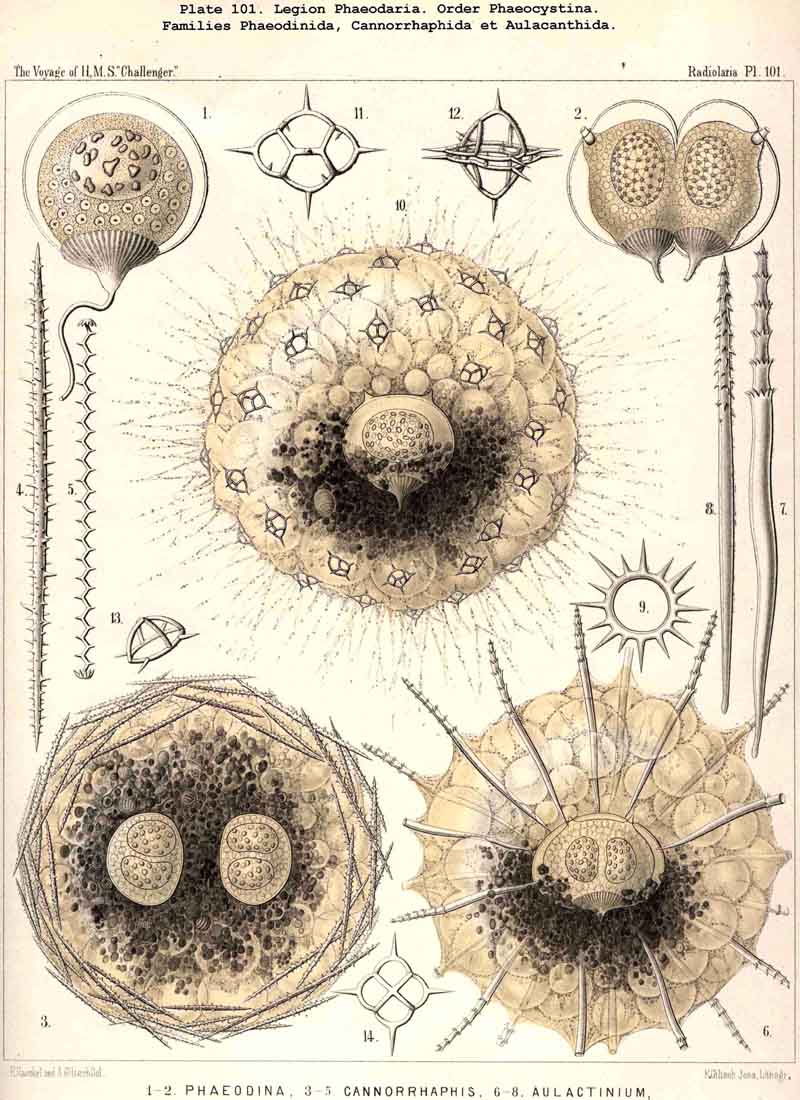
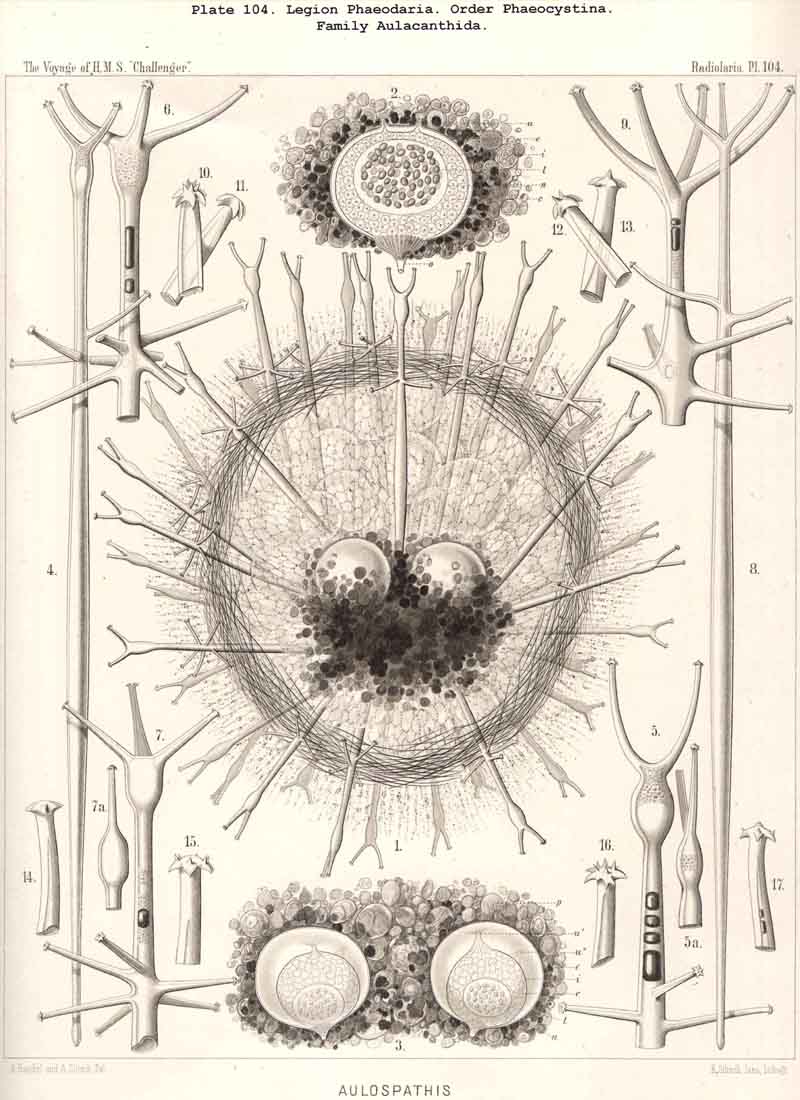
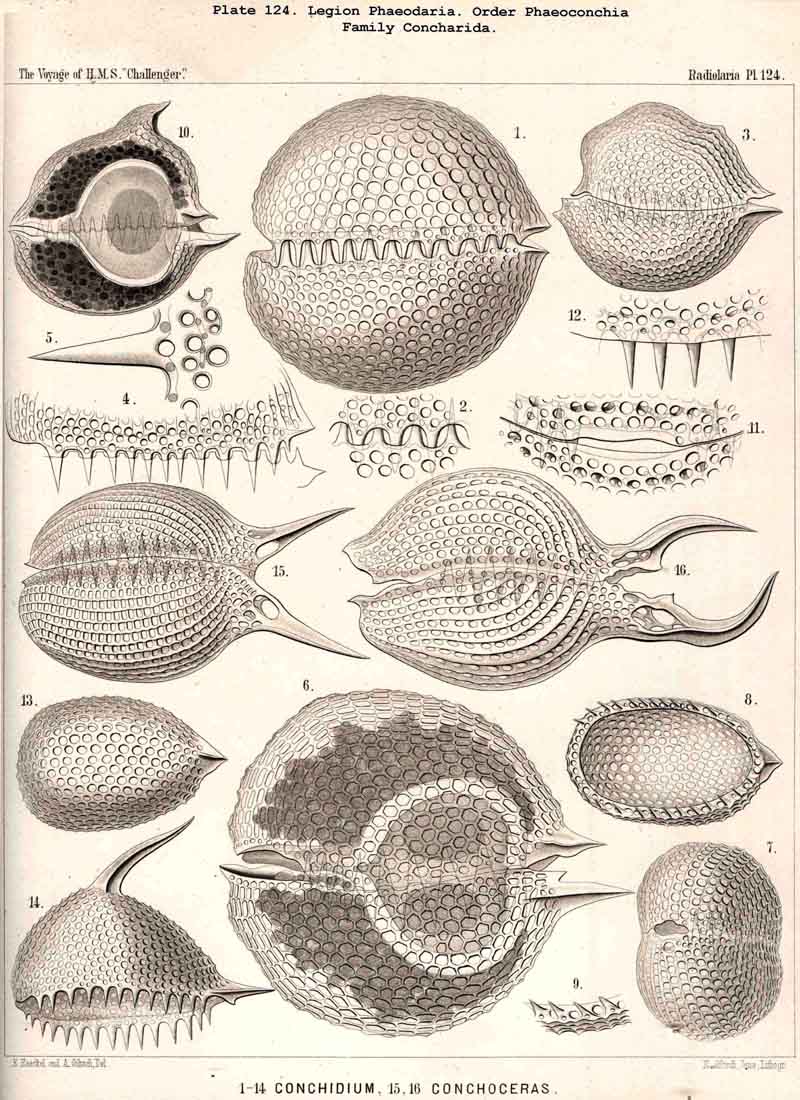
70. The Nucleus of the Cannopylea.-The nucleus presents the same remarkable structures in all species of the PHAEODARIA or CANNOPYLEA which have been examined, and closely resembles the germinal vesicle of an amphibian ovum, being a large spherical or spheroidal vesicle with numerous nucleoli. Its diameter usually amounts to half or two-thirds, sometimes even three-quarters, that of the central capsule. The vertical main axis of the latter is also that of the nucleus, which usually lies somewhat nearer to the aboral pole. The nucleus is generally rather more strongly compressed in the direction of the main axis than the capsule itself. The membrane of the vesicular nucleus is thin, but firm, and encloses a clear or finely granular mass of nuclein. The number and size of the contained nucleoli are variable even in one and the same species, and stand in inverse ratio to each other, an obvious result of the gradual process of division. Commonly from twenty to fifty roundish or spherical, strongly refracting nucleoli, are present; more rarely there are several hundred very small ones. Sometimes the nucleus is penetrated by fine trabeculae, in whose meshes lie the nucleoli (Pl. 101, fig. 2). In certain nuclei, which contained a few large nucleoli, these were of irregular form, probably the result of amoeboid movements (Pl. 101, fig. 1). In the formation of spores in the CANNOPYLEA, the nucleus apparently becomes dissolved, and its numerous nucleoli develop directly into the nuclei or mother-nuclei, which produce the nuclei of the flagellate spores. Further more, many PHAEODARIA seem to multiply by simple cell-division, since very commonly (especially in the Phaeocystina and Phaeoconchia) two large nuclei (right and left), may be met with in one central capsule; sometimes also a single large nucleus, in which a sagittal constriction marks the commencing division of the capsule (Pl. 101, figs. 2, 36; Pl. 104, fig. 3; Pl. 124, fig. 6, &c.).
The large nucleus of the PHAEODARIA was first described in my Monograph in 1862, in the case of Aulacantha (p. 263), Aulosphaera (p. 359), and Coelodendrum (p. 361), as a "large, spherical, thin walled endocyst," from 0.1 to 0.2 mm. in diameter. More detailed descriptions, especially with respect to the behaviour of the nucleoli were given by R. Hertwig in 1879 (L. N. 33, p. 97).
71. The Endoplasm or Intracapsular Protoplasm.-In all Radiolaria the intracapsular protoplasm, which, for the sake of brevity, may be termed "endoplasm," constitutes originally, and especially in the earliest stages, the only important content of the central capsule, except the nucleus. In certain SPUMELLARIA and NASSELLARIA, of simple structure and of small dimensions, this condition persists for a long period, and the endoplasm then appears as a homogeneous, colourless, turbid or finely granular, mucous, semi-solid mass, which cannot be distinguished from the ordinary undifferentiated proto plasm of young cells; no definite structure, and in particular, no fibrillar network, can be discovered in it even by the use of the customary reagents. In the great majority of the Radiolaria, however, this primitive homogeneous condition of the endoplasm is very transient, and it soon undergoes definite modifications, becoming differentiated into separate parts or producing new constituent contents. Such products of the internal protoplasm are in particular hyaline spheres (vacuoles and alveoles), oil-gbobules, pigment- bodies, crystals, &c. The most important of the differentiations which take place in the endoplasm is that into an internal, granular, medullary substance and an external, fibrillar, cortical substance; although the various legions behave somewhat differently in this respect (§§ 77-80).
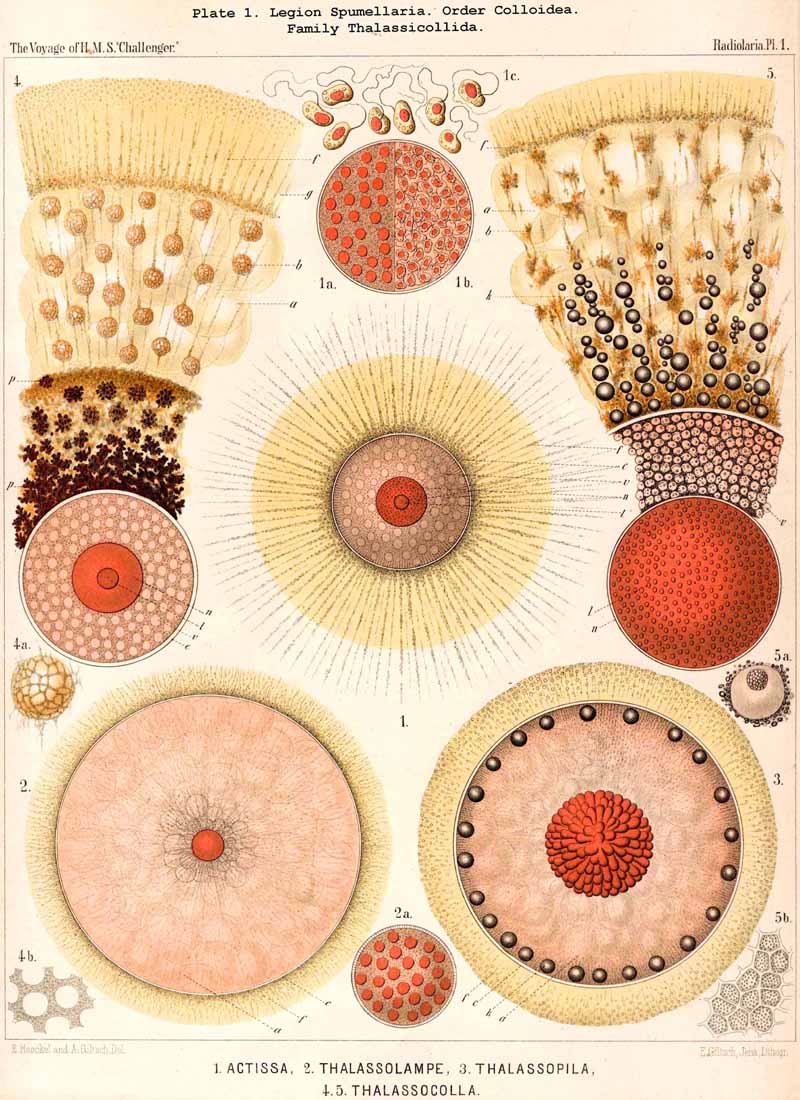
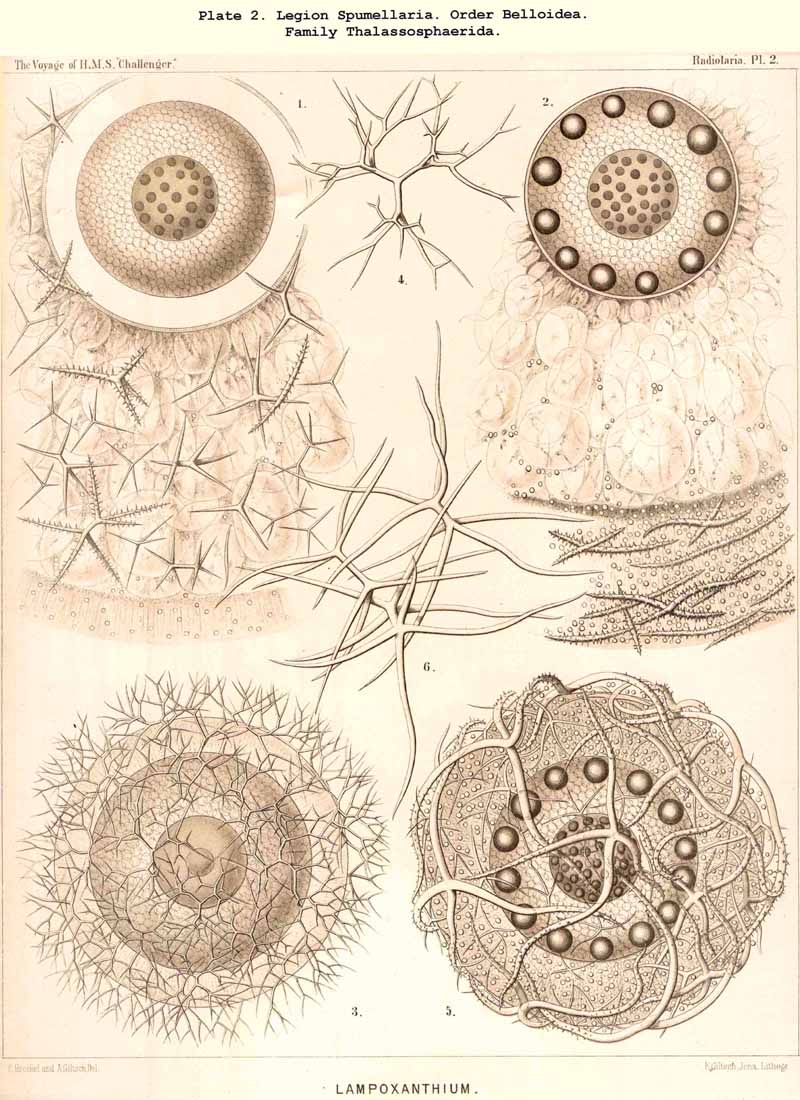
72. Intracapsular Hyaline Spheres.-The central capsule of very many Radiolaria contains in its endoplasm numerous spherical bodies of varying size, which consist of watery or albuminous fluid, and have previously been regarded as nuclei, or described as products of the internal protoplasm, under various names, such as "spherical transparent vesicles" (see note A, below), "albumen spheres" (see B), "gelatinous spheres" (see C), "alveolar cells" (see D), &c. Some of these spheres are perfectly transparent, structureless and of varying refractive power, producing the impression of drops of fluid; others contain various formed constituents, such as oil-globules, fat-granules, pigment granules, concretions, crystals, &c. From a morphological point of view they may all be divided into two categories, membraneless vacuoles and vesicular alveoles. The vacuoles are simple spherical drops of fluid or of gelatinous material, devoid of a special envelope, but immediately surrounded by the endoplasm. The alveoles, on the other hand, are true vesicles with a thin spherical envelope, enclosing a drop of fluid or jelly. This envelope is commonly very thin, homogeneous, and often scarcely discernible, so that in practice a sharp line of demarcation cannot be drawn between alveoles and vacuoles; the former are usually somewhat larger than the latter. The fact is, nevertheless, certain that the hyaline spheres, which may be isolated on rupturing the central capsule of many Radiolaria, in certain cases, particularly in large species, possess a clear, anatomically demonstrable membrane, whilst in others no such appearance is presented. It may be assumed that the vesicular alveoles are developed from the drop-like vacuoles by increase in size, and by the precipitation of a delicate envelope from the endoplasm. The character common to all these hyaline spheres, whether vacuoles or alveoles, is found in their aqueous, not adipose, constitution, and in their clear transparent appearance, which allows of no structure (the above-mentioned contained bodies excepted) being recognised. Their refractive power and consistency vary somewhat, and probably their chemical constitution still more. Sometimes they are strongly refractive and shining, and sometimes feebly refractive and pale; their consistency shows all intermediate stages between a thin fluid, which readily disappears in water, and a firm, insoluble jelly. As regards their chemical composition (which is probably very variable), the hyaline spheres may be best divided into two groups, the organic and inorganic. The inorganic hyaline spheres are simple drops of saline solution without any carbonaceous constituent; the organic, on the other hand, contain a small quantity of organic matter dissolved in the watery fluid, and may be either albuminous or gelatinous spheres. The formed contents which are commonly present are of very various natures, usually small fat- granules, more rarely larger fat-granules or pigment-granules, sometimes concretions or crystals. In many groups, especially among the large PHAEODARIA and Collodaria, the numerous hyaline spheres are remarkable for their equal size and even distribution throughout the endoplasm (Pl. 1, figs. 1, 4; Pl. 104, fig. 2, &c.). In some genera belonging to the Thalassicollida the alveoles are of enormous size (Pl. 1, figs. 2, 3); they then become flattened by mutual pressure into polyhedra and distend the central capsule to unusual dimensions (in Physematium and Thalassolampe 8 to 12 mm.).
A. The "spherical hyaline vesicles," which I described in my Monograph (1862, p. 71) as among the most important and constant contents of the central capsule, are partly vacuoles, partly homogeneous nuclei. Most recent investigators, Bütschli in particular (1882, L. N. 41), have pointed out and rightly criticised this confusion. The criticism might, however, have been more justly expressed by stating that, in the preparation of my Monograph (1859-1862), I did not make use of modern methods of demonstrating the nucleus by staining fluids, which were quite unknown at the time, and only discovered a decade later. In fact, without the aid of such reagents, it is quite impossible to distinguish between the various "spherical transparent vesicles," of which those found in the central capsule of the PHAEODARIA and many monozootic Collodaria are simple vacuoles lying in the endoplasm, whilst, on the other hand; those of the Polycyttaria and many other Radiolaria are true homogeneous nuclei. For not only are the general appearance of the small clear spheres, their refractive power. and regular distribution in the endoplasm quite similar, but they are also of much the same size, for the diameter ranges from 0.005 to 0.015 mm., being generally between 0.008 and 0.012 mm. In addition to this there is generally in each hyaline sphere a dark brightly shining granule, which, in the case of the vacuole, is simply a fat-granule, whilst in the case of the nucleus, it is a true nucleolus. The small hyaline spheres in the young uninucleate capsules of the Polycyttaria are simple vacuoles (Pl. 3, fig. 12), whilst in the ripe multinucleate capsules they are true nuclei (Pl. 3, figs 3, 8, 9), and it is quite impossible to discriminate between these two conditions without the use of reagents. This has been expressly recognised by R. Hertwig, who has the merit of having been the first to clearly distinguish, by the aid of staining fluids, between these two different constituents (1879, L. N. 33, p. 108).
B. The "albumen spheres," which were first observed by A. Schneider in 1858 in the common cosmopolitan Thalassicolla nucleata (L. N. 13, p. 40), and which appear to occur in only a few other Thalassicollida, are distinguished from the ordinary hyaline spheres of about the same size by their higher refractive power and by certain albuminoid reactions, especially the coagulation of a membranous envelope under the influence of certain reagents (see my Monograph, p. 250, and Hertwig, L. N. 26, 1876, p. 46). They often enclose various formed contents, and require further investigation.
C. The gelatinous spheres of various sizes, found in the endoplasm of the Radiolaria, agree in their reactions (especially in staining by certain reagents) with the common extracapsular jelly of the calymma, and are hence distinguishable both from, the true (coagulable) "albumen sphere," and from the ordinary watery vacuoles.
D. The alveoles, which are only accurately known in the case of certain large monozootic Collodaria, but which also seem to occur in the central capsule of other remarkably large Radiolaria, were described in my Monograph in the case of Thalassolampe margarodes and Physematium mülleri, under the name "intracapsular alveolar cells" (1862, pp. 77, 254, 257). They are not, however, true nucleated cells, and the body described as a nucleus is not such in reality. Nevertheless these large hyaline spheres do possess a special envelope, as I have recently convinced myself by the examination of ruptured central capsules of Thalassolampe maxima, Thalassopila cladococcus, and Physematium atlanticum (Pl. 1, figs 2, 3). The central capsule of these Collodaria becomes distended to most unusual dimensions (2 to 12 mm. in diameter) by the great development of these large hyaline vesicles, each of which measures from 0.1 to 0.5 mm. in diameter.
73. The Intracapsular Fat-Globules.-Fat is present in the central capsule of all Radiolaria in larger or smaller quantities, and generally appears in the form of very numerous, small, spherical granules, which are either distributed evenly in the endoplasm (as an emulsion) or enclosed in the vacuoles; the latter, in particular, is the case in most PHAEODARIA, perhaps generally. In this group each vacuole contains as a rule a single dark, shining fat-granule, and sometimes also an irregular bunch composed of from two to five or more granules. In addition to these small fat-granules (granula adiposa) which are always present, the central capsule of many Radiolaria contains also larger fat-gbobules (globuli adiposi). These appear to be generally wanting in the PHAEODARIA, and are on the whole rare in the ACANTHARIA; whilst, on the contrary, they are very common in the NASSELLARIA and SPUMELLARIA. The Polycyttaria or social Radiolaria are as a rule distinguished by the possession of a single large central oil-globule, which lies in the centre of the central capsule, and is on an average about one-third of it in diameter (Pl. 3, figs. 4, 5). This is absent, however, in those young capsules of the Polycyttaria in which the primary nucleus is centrally situated (Pl. 3, fig. 12). Those species of Polycyttaria whose central capsule reaches a considerable size, often enclose numerous oil-globules, and in Collophidium (species of Collozoum with an elongated cylindrical capsule, Pl. 3, figs. 1, 3) the axis of each capsule is occupied by a row of numerous oil-globules. In the monozootic SPUMELLARIA, in which the nucleus is always centrally situated, the large oil-globules are, of course, excentric, being in apposition to the inner surface of the capsule-membrane (Pl. 1, fig. 3; Pl. 2, figs. 2, 5). In the Discoidea the oil-globules, which are often present in large numbers, form elegant concentric rings around the central nucleus, and in those species with segmented arms, there are one or more transverse rows in each segment (Pl. 43, fig. 15). In the NASSELLARIA the number and distribution of the oil-globules are dependent upon the form of the central capsule. When this is simple, without lobes, and ovoid or conical, they generally lie in its aboral half above the podoconus (Pl. 51, figs. 5, 13; Pl. 97, fig. 1). When, on the contrary, the basal portion of the capsule sends out three or four dependent processes (as in the majority of the Cyrtoidea), a large globule may generally be seen in the swollen distal part of each conical or ovoid lobe (Pl. 53, fig. 19; Pl. 60, figs. 4-7). In many Stephoidea and Spyroidea, whose central capsule is separated into two lateral portions by the constriction corresponding to the sagittal ring, each of these contains either a single large globule or a group of small ones (Pl. 90, figs. 7, 10). These oil-globules are usually colourless and highly refractive; rarely they are yellow or brown, sometimes rose-coloured, or an intense blood-red (e.g., in Thalassophysa sanguinolenta) or even orange (in Physematium mülleri). In many SPUMELLARIA, and particularly in the Polycyttaria, an albuminous substratum may be recognised in them, which is sometimes disposed in layers, and after extraction of the fat presents the appearance of a laminated sphere. The physiological significance of the oil-globules is twofold; in the first place they tend to diminish the specific gravity of the organism; in the second they may be utilised as a reserve store of nutriment. In the latter respect they are of special importance in the process of spore-formation, each flagellate spore usually containing a fat-granule.
74. The Intracapsular Pigment-Bodies.-In the majority of Radiolaria when observed alive, the central capsule is coloured, only in the minority is it colourless. The colour is never diffuse, but always due to the formation of definite pigment-granules or vesicles, which are sometimes distributed evenly throughout the endoplasm, sometimes aggregated in the central or peripheral regions. Their form may be either spherical, irregularly rounded, or polyhedral. They vary much in dimensions, but in most cases are immeasurably small, and appear under a high magnifying power as fine dust; occasionally, however, their diameter may amount to from 0.001 to 0.005 or more. The chemical constitution of the intracapsular pigment is unknown in most Radiolaria, and is probably very various. In many instances the pigment-granules consist of fat, in others not. The commonest colours are yellow, red, and brown; violet and blue are rare, and green still rarer. Sometimes a definite tone of colour prevails throughout a whole group, and may then be attributed to inheritance, e.g., red is found in most Sphaeroidea, and blue in the Polycyttaria (see note A). One colour is almost always constant in the members of the same species. True pigment-cells, belonging to the Radiolarian organism, do not occur within the central capsule. The peculiar yellow cells which are found in the central capsule of many ACANTHARIA are symbiotic xanthellae (see § 76).
A. The number of Radiolaria whose pigment has been examined in the living state, is too small to allow of any general conclusions being drawn. Regarding the different colours known, see my Monograph, L. N. 16, p. 76.
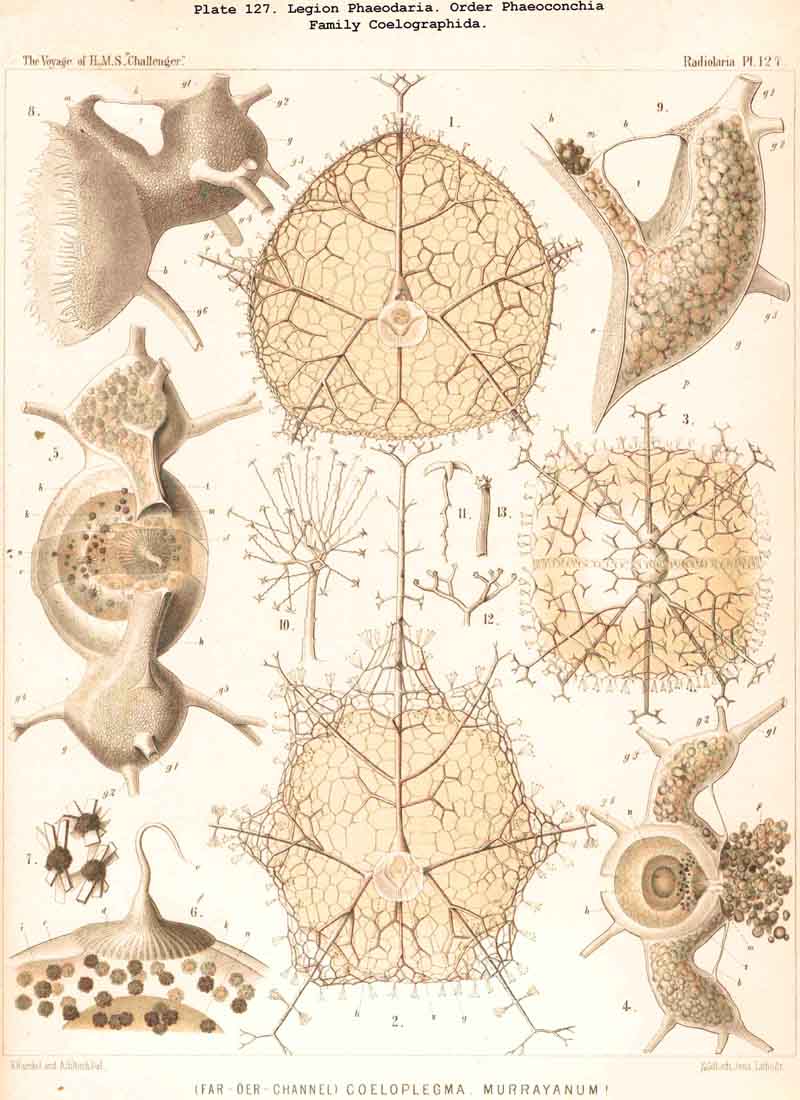
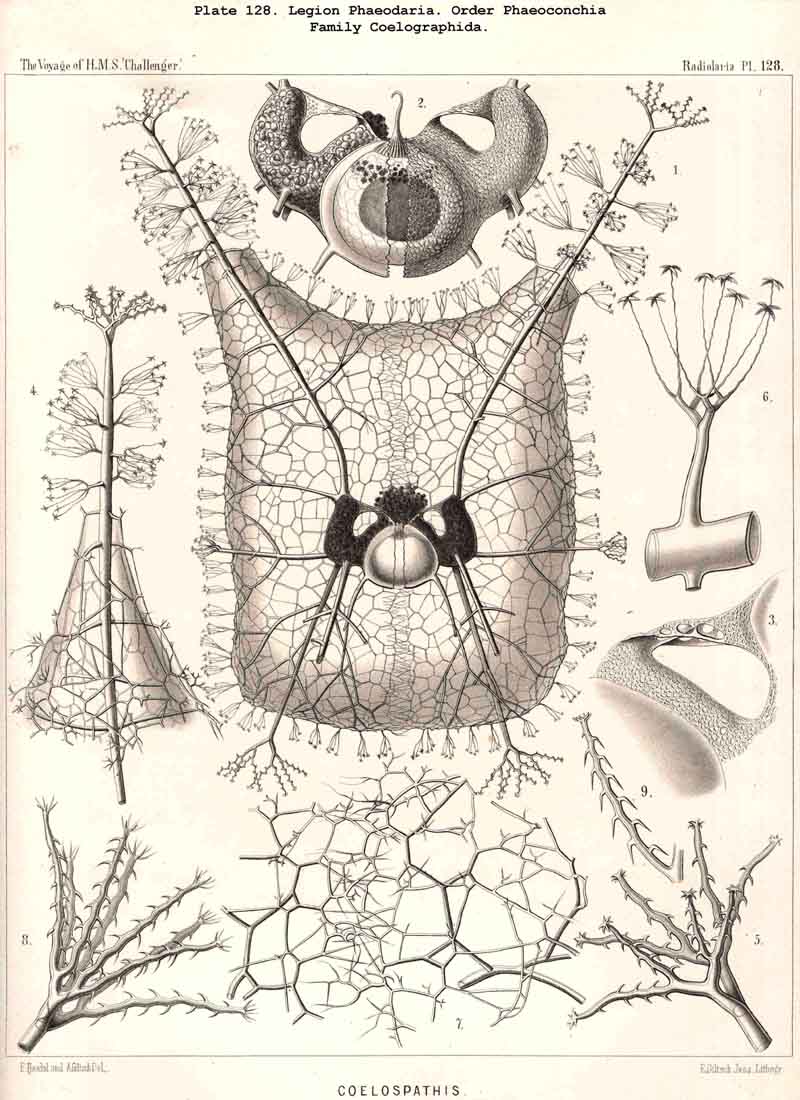
75. The Intracapsular Crystals.-The crystals found in the central capsule of many Radiolaria may be divided into two groups, of very different significance; small crystals, which are very widely distributed, and large crystals, which occur in only a few genera. The small crystals may also be termed "spore-crystals," since each swarm-spore often contains such a crystal. They are rod-like or spindle-shaped, and consist of an organic substance which probably serves as a reserve of nutriment for the developing spores. Such spore-crystals have been observed in numerous SPUMELLARIA and ACANTHARIA belonging to various families, and are probably present throughout the two legions which make up the Porulosa. On the other hand, they have not been noticed in the Osculosa (NASSELLARIA aiid PHAEODARIA), the few swarm-spores belonging to these groups which have been observed not exhibiting any crystals. The large crystals, which occur in small numbers in the endoplasm, have hitherto only been observed in a few species of SPUMELLARIA, belonging to the Polycyttaria. They were first noticed in the common Collosphaera huxleyi, and regarded as coelestin. They are also found in the central capsule of many other Collosphaerida, e.g., Buccinosphaera (Pl. 5, figs. 11, 12). Crystal-masses, crystal-sheaves, or spherical masses of radiating acicular crystals are enclosed in the vacuoles or "albumen globules" of Thalassicola nucleata and other Thalassicollida, as well as in the central capsule of Caelographis and some other PHAEODARIA (Pl. 127, figs. 4-7). All these large crystals are probably to be regarded as excretory products.
75A. The Intracapsular Concrements.Concretions, either mineral or organic, of varying form and constitution, are to be found in the endoplasm of Radiolaria belonging to very different families. They are most abundant and multiform in Thalassicolla nucleata, being usually circular or elliptical discs, which are concentrically laminated and highly refractive, resembling starch-grains. Among them twin forms may frequently be observed, as though the concrements were in process of division (see note A). Similar amyloid concretions are to be seen in the central capsule of different SPUMELLARIA and NASSELLARIA, e.g., in Cephalospyris triangulata (Pl. 96, fig. 28). Violin-shaped, highly refractive concrements have been observed in the central capsule of numerous SPUMELLARIA, NASSELLARIA, and ACANTHARIA, e.g., Thalassosphaera, Spongosphaera, Plegmosphaera, Cyrtocalpis, Peripyramis, Botryocella, &c. (see note B). The chemical constitution of these concrements is insufficiently known.
A. The amyloid concretions of Thalassicolla nucleata have been described in detail in my Monograph (pp. 80, 250, Taf. iii. figs. 2, 3), and by R. Hertwig in the Histologie der Radiolarien (1876, p. 47, Taf. iii. figs. 9-13).
B. The violin-shaped concretions of Thalassosphaera bifurca have been figured in my Monograph (pp. 80, 261, Taf. xii. fig. 1).
76. The Intracapsular Xanthellae.-The xanthellae, zooxanthellae, or symbiotic "yellow cells" are found within the central capsule only in the ACANTHARIA, whilst in other Radiolaria they only occur in the extracapsulum. They are most frequent in the Acanthometra, rarer in the Acanthophracta, but even in the former they are often wanting. Their number is very variable, but usually small, from ten to thirty in one capsule. They lie for the most part immediately below the capsule membrane, in the cortical layer of the. endoplasm. The form of the yellow cells is either spherical or ellipsoidal, often also spheroidal or even lentiform. The diameter varies from 0.01 to 0 .03 mm. They possess a distinct membrane and an excentric nucleus, and contain numerous yellow pigment-granules in the endoplasm. This yellow pigment dissolves in mineral acids to form a green fluid, and in other respects also behaves somewhat differ ently from the yellow pigment in the extracapsular yellow cells of the SPUMELLARIA and NASSELLARIA. In both cases, however, the xanthellae are not integral portions of the organism, but unicellular algae, living as parasites or symbiontes in the body.
A. The yellow cells in the central capsule of the ACANTHARIA were first observed by Joh. Müller (L. N. 12, pp. 14, 47). In my Monograph I described them at greater length, and indicated their differences from the extracapsular yellow cells of other Radiolaria (L. N. 16, pp. 77, 86). Since then, R. Hertwig has demonstrated their cellular nature (L. N. 33, pp. 12, 113), and still more recently Brandt has given further accurate information regarding their occurrence, constitution, and physiological significance (L. N. 39, ii. Art., p. 235, figs. 62-73).
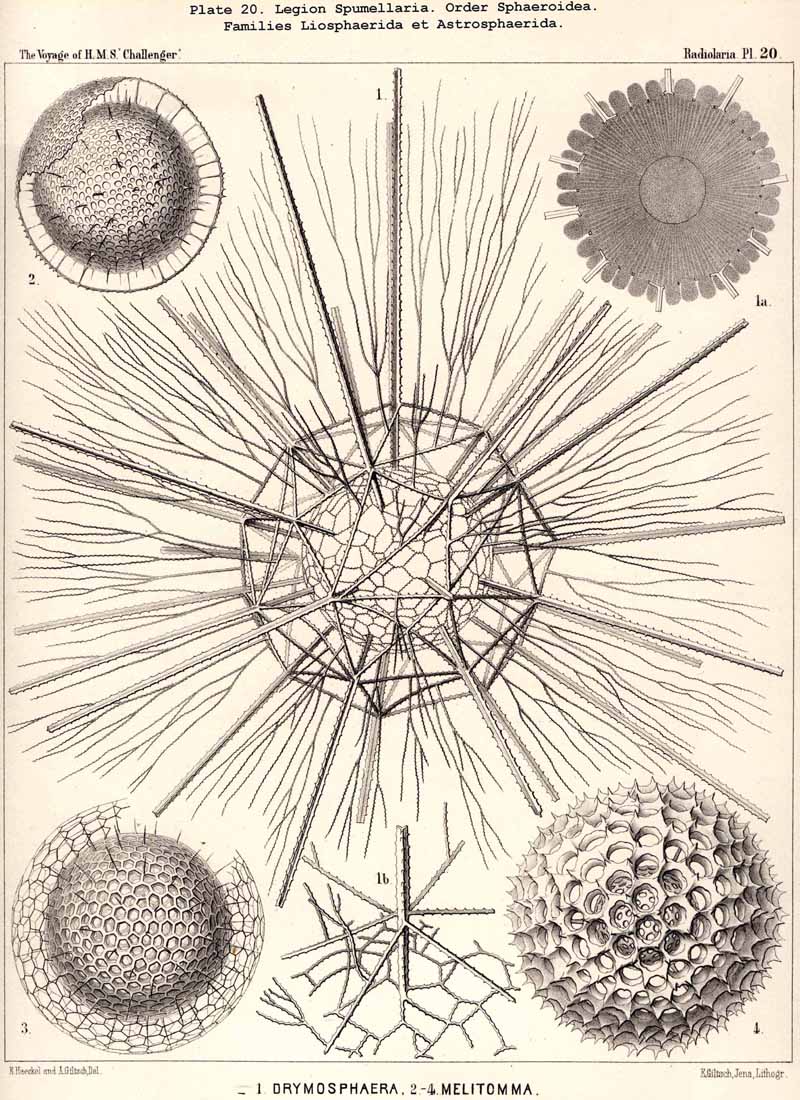
77. The Endoplasm of the Peripylea.-The intracapsular protoplasm of the SPUMELLARIA or PERIPYLEA is usually distinguished by a more or less complete radial arrangement, which does not occur in the same form in other Radiolaria; it may be regarded as characteristic of this legion, for it probably occurs in all the species at some period of life or other, and stands in a direct causal relationship with the typical structure of the capsule-membrane in all the "PERIPYLEA" (see note A). For as this is commonly perforated by very numerous pores distributed at equal intervals over the whole surface of the capsule, and since a communication between the intra- and extracapsular sarcode takes place through these, the radiate structure of the endoplasm may be readily explained as due to the influence of radial currents which take place continuously or intermittently in the endoplasm. This radiate structure is most obvious when the endoplasm contains no secondary products or only an insignificant amount of these, and thus appears colourless and almost homogeneous, or only finely granular. Under these circumstances, an optical section of the central capsule usually reveals a distinct radial striation; numerous narrow, straight, dark streaks alternating regularly with still narrower clear ones; the latter consist of homogeneous, the former of more or less granular protoplasm (Pl. 20,fig. 1a). Often there may be distinguished in each darker streak a single straight row of strongly refracting (fat?) granules, sometimes several such rows. Occasionally the whole endoplasm becomes divided up into a number of large "radial wedges," club-shaped, conical or pyramidal masses of granular protoplasm, separated by clear divisions of hyaline plasma (e.g., in Actissa radiata, p. 14, where in the optical section of the central capsule, between the membrane and the nucleus, twenty-five dark radial wedges of equal size were separated by thick clear partitions of hyaline protoplasm). In the majority of the SPUMELLARIA this radial striation is partially or entirely concealed by the formation of pigment or of other products. Very often it is only visible in the cortical layer, which lies immediately below the capsule-membrane (Pl. 1, figs. ii, 3). The remarkable "centripetal cones" which characterise the Thalassicollid genus Physematium, and were formerly described as "centripetal cell-groups," are probably a special development of these cortical radial wedges; they are conical cortical bodies, regularly distributed on the inner surface of the membrane of the central capsule, and disposed with the apex turned towards the centre (see note B). More rarely than in the cortical layer, a similar radial structure is to be found in the innermost medullary layer immediately surrounding the nucleus. Here the endoplasm sometimes breaks up into fine radial threads, which are anatomically separable and hang down from the free nucleus as thin processes (see note C). In some cases it is also possible to isolate radial rods from the cortical layer of teased out central capsules.
A. The radial structure of the endoplasm was first described in my Monograph (1862, p. 74), though R. Hertwig (1879, p. 112) was the first to indicate its typical significance in the case of the PERIPYLEA, and to demonstrate its causal relation with the radial currents in the central capsule of this legion. More recent investigations have led me to the conviction that this phenomenon is more widespread, and often more strongly developed, than was formerly imagined, and that it is probably one of the typical characters of all SPUMELLARIA (at least of the Monozoa).
B. The centripetal cones of Physematium, which have hitherto been known only in these colossal Thalassosphaerida, were fully described in my Monograph under the name "conical centripetal cell-groups"; by their first discoverer, A. Schneider (L. N. 13), they were termed " nests, " and compared with the "nests" (central capsules) of the Polycyttaria. In the Physematium mülleri of the Meditteranean (hitherto only observed by Schneider and myself at Messina) it appeared as though each centripetal cone were composed of a group of from three to nine (usually four or five) slender wedge-shaped cells, whose common centripetal apex was produced into a radial thread of sarcode (L. N. 16, p. 258, Taf. iii. fig. 7). Since then (1866) I have observed at Lanzerote, in the Canary Islands, a nearly related form, which I take to be Physematium atlanticum, Meyen. In this, however, the "centripetal cell-groups" were wanting, and the whole cortical layer of the endoplasm was cleft into numerous radial portions, each enclosing a nucleus (probably the mother-cells of flagellate spores, see p. 35).
C. The radial fibres of the medullary endoplasm which cling to an extracted
nucleus have been observed by Hertwig in certain Sphaeroidea (Diplosphaera,
Arachnosphaera) (L. N. 33, p. 40).
78. The Endoplasm of the Actipylea.-The intracapsular protoplasm of the ACANTHARIA or ACTIPYLEA is often distinguished by a partial or complete radial arrangement like that of the PERIPYLEA, but differing in the number, size, form, and distribution of the radial portions into which the endoplasm is differentiated. For since the pores of the capsule membrane are distributed at equal distances all over the surface in the SPUMELLARIA, whilst in the ACANTHARIA they are arranged in definite groups, and since the number and arrangement of the pores has a direct influence upon the internal currents of the endoplasm, it follows that the radial structure in the latter legion must be very different from that in the former. In addition to this there must not be forgotten the important influence which the early centrogenous formation of the skeletal rods exercises upon the disposition and. growth of the intracapsular structures. Hence the endoplasm of the ACANTHARIA does not separate into innumerable thin, closely packed radial wedges or cortical radial rods, but into a small number of large pyramidal portions between which run the radially disposed heterogeneous portions of the contents of the capsule, viz., the radial bars of acanthin and the peculiar intra capsular "axial threads." As a direct consequence of the regular disposition of these heterogeneous radial portions, which is often characteristic of the various families of the ACANTHARIA, a corresponding differentiation of the endoplasm is brought about; it divides into a number of conical or pyramidal portions (radial pyramids), whose bases rest upon the capsule-membrane and whose apices are directed towards the centre of the capsule (the central star of the skeleton). These radial pyramids are, however, but rarely visible, being usually more or less concealed by a dark pigment.
The differentiations of the endoplasm in the central capsule of the ACTIPYLEA have been but little investigated, but they appear to vary somewhat in the different groups of this legion. In all ACANTHARIA in which the twenty radial bars are regularly arranged according to the Müllerian law (see p. 717) and in which axial threads constant in number and disposition run between them from the central star to the capsule-membrane, it obviously follows that the endoplasm must be divided into more or less distinct radial pyramids, and this must be the case whether these take the form of continuous tracts or of actually separable portions. The regular polygonal figures, often seen on the surface of the central capsule (with special distinctness in Acanthometron elasticuim and Acanthometron pellucidum) separated by a network of granular threads, are the bases of such radial pyramids (see Hertwig, L. N. 43, p. 12, Taf. i. figs. 1-7).
79. The Endoplasm of the Monopylea.-The intracapsubar protoplasm of the NASSELLARIA or MONOPYLEA is distinguished from that of any of the other three legions by the development of a quite peculiar fibrillar structure, the axial "pseudopodial cone," which may shortly be termed the "podoconus" (foot-cone). Since this is in direct correlation with the peculiar structure of the capsular opening, the large "porochora," which is situated at the basal pole of the main axis, it is quite as characteristic of the legion as the latter itself (see note A). The podoconus is primitively a vertical regular cone, whose circular base occupies the horizontal porochora or "basal porous area" of the central capsule, while its vertical axis coincides with that of the latter. The apex of the cone, usually somewhat rounded off, is therefore directed towards the aboral or apical pole of the central capsule and separated from it by a larger or smaller interval. In this interval the nucleus originally lies (as in Pl. 51, fig. 13; Pl. 98, fig. 13); but it is usually displaced subsequently and lies excentrically. The cone is of very variable height; on an average its vertical height is about equal to the diameter of its horizontal base; these dimensions are, however, dependent upon the form of the central capsule; the height being greater in slender ovoid or conical capsules, and less in depressed sphaeroidal or discoidal ones, than the diameter of the base. The podoconus consists of differentiated endoplasm, which becomes more deeply stained by carmine and offers greater resistance to solvents than the surrounding finely granular protoplasm. The apex, especially, becomes very intensely stained. It always exhibits a very characteristic fine but distinct striation, numerous straight radial lines diverging from the apex of the cone towards the base. The number of these striae appears to correspond with that of the vertical rods in the porochora, and each of these latter stands apparently in direct communication with the basal end of an apical stria (§ 59). These threads are probably differentiated constant contractile threads of endoplasm, or even myophanes, comparable with the contractile cortical threads of the CANNOPYLEA and the permanent axial threads of the ACTIPYLEA. The numerous modifications, undergone by the form and contents of the central capsule in the different groups of MONOPYLEA, especially those due to the formation of the skeleton, are not without influence upon the podoconus. The most important divergencies from the above described primary form are the following :- (1) The vertical axial cone becomes oblique, its axis inclining in the sagittal plane and approaching either the dorsal or the ventral wall of the capsule; the cause of this appears to be usually the excentric development of the growing nucleus or the formation of a large oil-globule. (2) The smooth mantle of the podoconus becomes divided by three longitudinal furrows into three equal pro minent ridges, which correspond to three circular lobes in the porochora; the cause of this basal triradial lobular formation lies probably in the triradial development of the skeleton in many NASSELLARIA or in the cortinar structure of the collar septum. (3) The simple podoconus splits into three or four elongated lobes, which eventually become almost completely separated and correspond to the lobes of the central capsule, in the axial wall of which they lie as longitudinally striated bands. The behaviour of these bands justifies the hypothesis that the podoconus is a muscular differentiated portion of the endoplasm and is composed of myophane fibrillae, whose contraction determines the opening of the central capsule.
A. The podoconus of the MONOPYLEA was first described by R. Hertwig in 1879, and recognised as a characteristic component of the central capsule in the most various groups of this legion (in Plectoidea, Stephoidea, Spyroidea, and Cyrtoidea; see his figures, loc. cit., Taf. vii., viii., and the description, pp. 71, 73, 83, 106). Hertwig called it the "pseudopodial cone," and regarded it as a conical process of the capsule-membrane, which is developed from this latter and projects from the porous area into the interior of the central capsule; "it is penetrated by fine canals which arise at the apex of the cone, diverge towards the base, and terminate there in the rods of the pseudopodial area. The intracapsular protoplasm penetrates at the apex of the pseudopodial cone into its fine canals, runs along them and emerges from the rods of the porous area in the form of slender threads" (loc. cit., p. 19). I cannot agree with this view of Hertwig, although I have been able to confirm the accuracy of his description by my own observations upon numerous excellently stained and preserved preparations in the Challenger collection. As I have proved by numerous teased out preparations, and as Hertwig himself correctly states, "the cone is more readily detached from the membrane than from the protoplasm, when the capsule is teased" (loc. cit., p. 73). Hence I regard the podoconus not as a differentiated portion of the capsule-membrane but as endoplasm, and believe that it is composed of myophanes or "contractile muscular fibrils" in the same manner as the cortical layer of the CANNOPYLEA. Probably the contraction of these fibrils serves to raise the opercular rods and hence to allow the exit of the endoplasm through the pores which lie between these opercular rhabdillae (compare § 59).
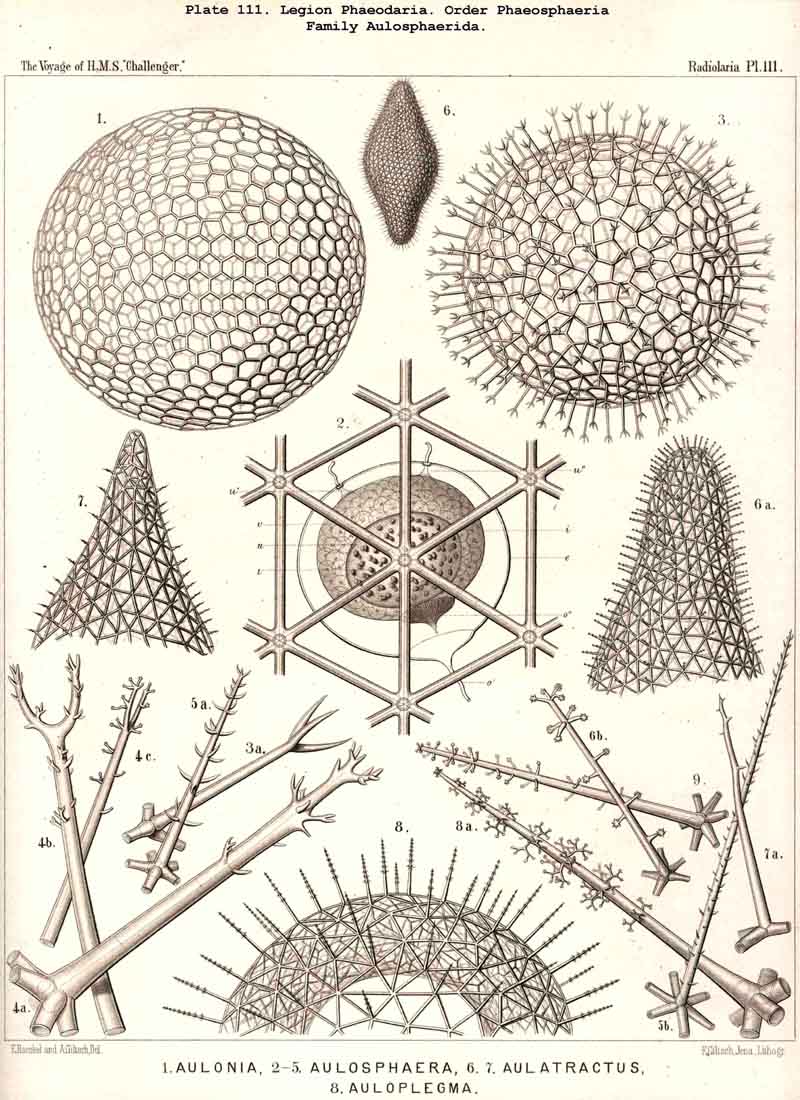
80. The Endoplasm of the Cannopylea.-The intracapsular protoplasm of the PHAEODARIA or CANNOPYLEA is distinguished from that of the other three legions by several characteristic peculiarities, which are very important, since they stand in causal relation to the typical structure of the capsule-membrane and in particular of its remarkable aperture. In the case of many and perhaps of all PHAEODARIA the endoplasm is differentiated into a granular medullary and a thin fibrillar cortical layer, the former of which usually encloses numerous small vacuoles, while the latter contains muscular fibrillae. In the voluminous central capsule of large PHAEODARIA the whole cortical layer of the endoplasm, which lies immediately below the delicate inner capsule-membrane, sometimes appears delicately and regularly striated, and most distinctly so under the apertures, towards the centre of each of which the dark striae are radially directed (see note A, below). These striae are probably contractile muscular fibrillae or "myophanes," by whose contraction the openings are voluntarily widened. In the Tripylea this fibrillar star is much more strongly developed under the astropyle (the main opening) than under the parapylae (or accessory openings); and probably the peculiar radial structure of the operculum of the former is due to the stronger development of these radial fibrils (being their impression). In many PHAEODARIA, indeed, the fine myophane fibrils are only visible under the apertures, whilst in others they form a continuous fibrillar cortical layer on the whole inner surface of the inner capsule-membrane; the fine fibrillae run meridionally from one pole of the main axis to the other; perhaps the whole central capsule may change its form in consequence of their contractions. The medullary portion of the endoplasm, which lies below this thin cortical layer, is usually finely' granular in the PHAEODARIA, and permeated by numerous spherical vacuoles, which are noteworthy from their equal size and regular distribution. Each clear vacuode usually contains a dark shining fat-granule, more rarely a group of such granules (see note B). Compare § 60, and Pl. 101, figs. 1-3; Pl. 104, figs. 1, 2; Pl. 111, fig. 2; Pl. 128, fig. 2, &c.
A. The fine fibrillae in the cortical layer of the endoplasm were first described by Hertwig in 1879 (L. N. 33, p. 98, Taf. x. figs. 6-10). He found them, however, only below the three openings in the capsule of the Tripylea, where they form three stellate groups of fibrils. I find them very clearly shown, and with especial distinctness, under the astropyle in most PHAEODARIA of which I have had the opportunity of examining well-stained and preserved central capsules. In many cases, also, the striation is not confined to the apertures, but spreads over the whole cortical layer. Perhaps this constitutes in all PHAEODARIA a thin myophane-sheet, whose contractile fibrils run from one pole of the main axis to the other and cause by their contraction changes in the form of the spheroidal central capsule.
B. The granular medullary portion of the endoplasm of the PHAEODARIA, with its numerous clear spherical vacuoles, was first described in my Monograph (1862), in the case of Aulacantha (p. 263), Aulosphaera (p. 359), and Coelodendrum (p. 361) as a "finely granular, mucous substance (intracapsular sarcode), packed more or less closely with clear spherical vesicles from 0.005 to 0.015 mm. in diameter, each of which contains one or two, rarely three, dark shining granules." That these clear spheres are true vacuoles was first clearly proved by Hertwig (L. N. 33, p. 98). As a rule all the vacuoles of the same central capsule are of equal size (generally from 0.008 to 0.012 mm. in diameter), and are distributed at equal intervals throughout the finely granular endoplasm.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}