{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

vel Actipylea, vel Acanthometrea (Pls. 129-140).
Acantharia, Haeckel, 1881.
Actipylea, Haeckel, 1882.
Acanthometrea, Hertwig, 1879.
Panacantha, Haeckel, 1878.
Definition.-Radiolaria with simple membrane bounding the central capsule, which is everywhere perforated by innumerable fine pores (disposed either equally or symmetrically). Extracapsulum without phaeodium. Skeleton centrogenous (its growth proceeding from the centre), acanthinic (organic, not siliceous). Fundamental form originally spherical.
The legion ACANTHARIA vel ACTIPYLEA, to the extent here defined, was constituted by me, 1878, in my Protistenreich (p. 102) under the name "PANACANTHA." A more accurate definition of this group was given in 1879 by Hertwig under the name ACANTHOMETREA. Both names were replaced by me, 1881, in my Prodromus (pp. 421, 465) by the more convenient name ACANTHARIA. This legion comprises all those Radiolaria which were first described by Johannes Müller, 1858, as Acanthometrae, and also an important part of his Haliomma.In my Monograph (1862, pp. 371-424) I disposed them in three families, Acanthometrida, Diploconida, and Dorataspida.
Although the number of genera and species in this legion is much increased by the rich collection of the Challenger, we can divide all ACANTHARIA into two different orders: Acanthometra (without complete lattice-shell) and Acanthophracta (provided with a complete lattice-shell).
The ACANTHARIA agree with the SPUMELLARIA in the structure of the simple capsule-membrane, which is perforated by numerous small pores (but constantly devoid of the large main opening, which the NASSELLARIA and PHAEODARIA possess, being hence united as "Merotrypasta"). We can therefore unite both former legions as "Holotrypasta" (compare above, pp. 5, 6); but in many ACANTHARIA (if not in all?) the numerous small pores of the capsule-membrane exhibit a certain peculiar arrangement not observed in the SPUMELLARIA; therefore the latter can be regarded as true " Peripylea" in opposition to the former as "Actipylea."
The peculiar main character of all ACTIPYLEA or ACANTHARIA is determined by the chemical constitution of their skeleton, which is not silex, but a peculiar organic substance, called by me in 1862 "acanthin" (Monogr. d. Radiol., pp. 30, 32). In all other Radioharia the skeleton is composed of silex or of a silicate. But besides this chemical difference, an important morphological character of the skeleton also separates the ACANTHARIA from all other Radiolaria: in the latter the skeleton is never centrogenous or arising from the centre of the capsule; in strict opposition to this general fact the skeleton of all ACANTHARIA is centrogenous, composed of radial spines, which arise from the central point of the capsule and pierce its membrane. These characteristic "radial spines of acanthin," arising from the centre, are never hollow (as formerly was supposed), but constantly solid. Their form is extremely variable, and most important for the distinction of genera and species; but more interesting from a general point of view is their peculiar arrangement or disposition.
The regular disposition of twenty radial spines has general value almost for all ACANTHARIA, with the exception only of the small group of Actinelida. In this latter group the number of radial spines is either more or less than twenty, and their disposition is either quite irregular or follows a peculiar rule. The number of individuals of these Actinelida, compared with that of the other ACANTHARIA, may be scarcely 1 per cent., whilst the latter have more than 99 per cent. ; the number of observed species is in the former about 5 per cent., in the latter about 95 per cent. Nevertheless the small group of Actinelida is very important, being probably the ancestral group from which all other ACANTHARIA have been phylogenetically derived. These other ACANTHARIA, with twenty regularly disposed radial spines, represent the two large groups of Acanthonida and Acanthophracta. For short and clear distinction of these two groups of ACANTHARIA, we will call the Actinelida (with irregular number and disposition of radial spines) Adelacantha, in opposition to the Icosacantha (Acanthonida and Acanthophracta), which all possess twenty regularly disposed radial spines.
Johannes Müller, the great zoologist, to whom we are indebted for the first detection and accurate knowledge of the Acanthometra, already recognised the regularity in the peculiar disposition of their twenty radial spines (Abhandh. d. k. Akad. d. Wiss. Berlin, 1858, pp. 12, 37). In honour of my great master I have called this regular disposition the "Müllerian law of spine disposition," and have given a full explanation of it in my Monograph (1862, pp. 40-45, 371, 372). With regard to its general value for all Icosacantha (Acanthonida and Acanthophracta), we might also call this pro-morphological Müllerian law "the Icosacanthan law."
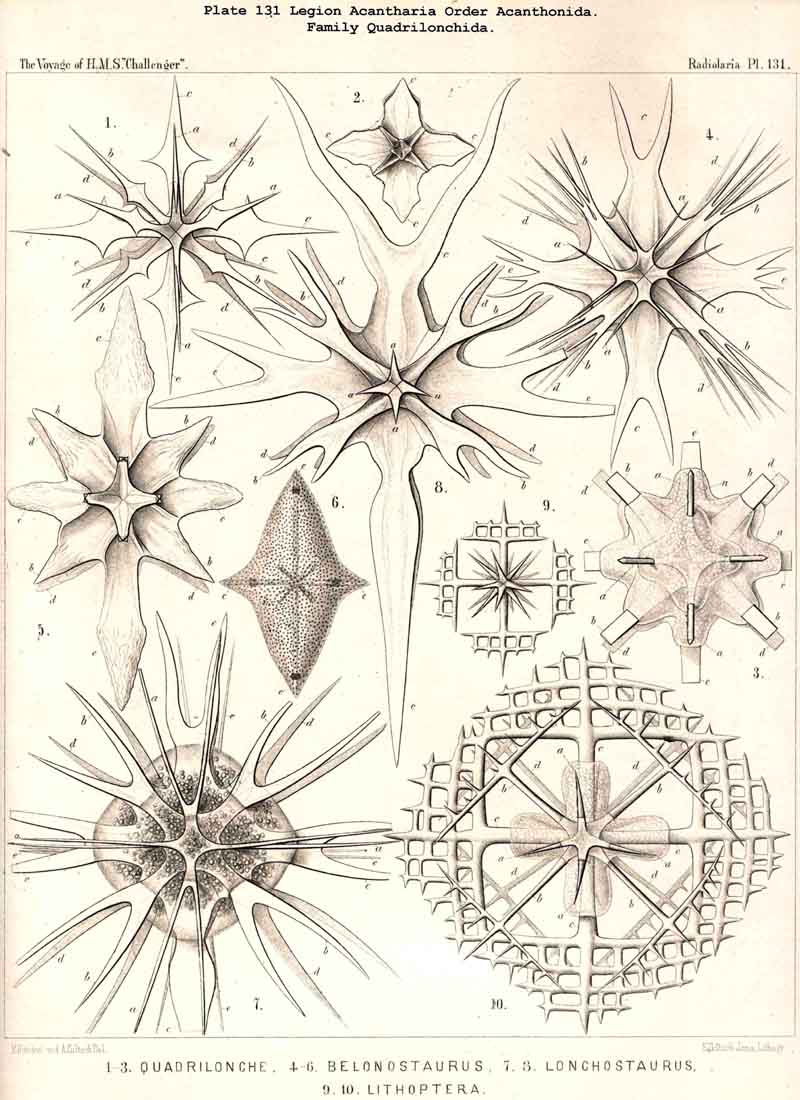
In 1862 I had already given the following precise definition of this "Icosacanthan law" (loc. cit., p. 40) :-" Between two poles of a spineless axis are regularly disposed five parallel zones, each with four radial spines; the four spines of each zone are equidistant one from another, and also equidistant from each pole; and the four spines of each zone are so alternating with those of each neighbouring zone, that all twenty spines together lie in four meridian planes, which intersect one another at an angle of 45°." For the clear conception of this remarkable Müllerian law, and for the complete understanding of its high value for the complicated morphology of all Icosacantha, it is the most profitable way to retain constantly in mind for comparison the figure of a terrestrial globe with its axis and zones. The axis of the globe is the spineless axis of all Icosacantha, around which all twenty spines are symmetrically disposed; it is perpendicular to the bisecting equatorial plane, in which lies the middle of the five parallel zones; therefore the, four spines, crossed perpendicularly in this equatorial plane, are called the equatorial spines (c1 to c4 in the figures of Pls. 131 -140); often, and mainly in the family Quadrilonchida (Pl. 131), these four equatorial spines are much larger or of a peculiar form, different from that of the sixteen other spines. Each pair of the four equatorial spines lies in one equatorial axis, and this latter is perpendicular to the crossing axis in which lies the other pair of opposite spines. We may regard these two equatorial diameters perpendicular one to another and to the spineless axis, as the two perradial axes or primary axes. Correspondingly the two meridian planes, which are determined by one perradial axis and the spineless axis, may be called the two primary or perradial meridian planes.
The globe is divided by the equatorial plane into two equal halves, the northern and the southern hemisphere. In each hemisphere there are disposed quite symmetrically eight radial spines, the distal ends of which fall in two parallel circles, a larger tropical circle (nearer to the equator) and a smaller polar circle (nearer to the pole of the spineless axis). Therefore we call the four spines of the former the "tropical spines and the four spines of the latter the "polar spines." The angle between the former and the equatorial plane is about 30°, the angle between the hatter and that plane about 60°.
The eight polar spines (four northern and four southern) lie in the same two meridian planes as the four, equatorial spines. Therefore in each of these two perradial planes lie six radial spines, opposite in pairs; two equatorial and four polar spines. Commonly all eight polar spines are of the same size and form; and often they are also equal to the eight tropical spines; but in some cases (e.g., in some species of Quadrilonchida) they are much smaller than the twelve other spines, and sometimes even rudimentary. In all figures of the Pls. 131 -140 (and also in my Monograph, 1862, Taf. xv.-xxii.) the polar spines of the northern circle are marked by the characters a1 to a4, the polar spines of the southern circle by the characters e1 to e4. In the first perradial meridian plane lie a1 and a3, e1 and e3, in the second a2 and a4, e2 and e4.
The eight tropical spines lie between the eight polar and the four equatorial spines, four in each hemisphere; their distal points fall in two parallel circles, which correspond exactly to the two tropics of the globe. Therefore the four northern tropical spines may be called "canceral spines" (as their ends fall in the Tropic of Cancer) and the four southern corresponding]y "capricornal spines" (as their points lie in the Tropic of the Capricorn). In the figures of the Pls. 131 -140 (as well as in my Monograph, 1862, Taf. xv.-xxii.) the four northern or canceral spines are marked by the characters b1 to b4, and the four southern or capricornal spines by the characters d1 to d4. Also the eight tropical spines lie (crossed in pairs) in two meridian planes; they do not lie, however, in those perradial planes, in which are placed the twelve other spines; but in two different meridian planes, crossing the former at angles of 45°; we call these the "secondary" or "interradial" meridian planes. Each of these planes is determined by the spineless axis and by two crossed inter-radial or secondary axes; in each of the latter lie two opposite tropical spines. In the first interradial meridian plane lie b1 and b3, d1 and d3, in the second b2 and b4, d2 and d4.
It is a most interesting and important fact, that in all Icosacantha (Acanthonida and Acanathophracta) this regular disposition of the twenty spines (in five parallel zones and four meridian planes) becomes constantly preserved by heredity, whilst the form and size of the different spines are extremely varied by adaptation.
Only in a minority of the Icosacantha are all twenty spines perfectly equal or nearly equal in size and form; and then it is often very difficult to distinguish the different zones in their disposition. But in far the greater part the size or the form of the twenty spines becomes different in different zones; and then we can commonly distinguish easily the five different zones. Firstly, in all Quadrilonchida and Dorataspida, the four equatorial are distinguished from the sixteen other spines either by form or by size, and often in a very remarkable degree. As soon as these four principal spines are recognised, it is easy to determine also the sixteen others; for the eight polar spines lie in the same two (perradial) meridian planes as the former, whilst the eight tropical spines lie in two different (interradial) meridian planes, intersecting the two former at angles of 45°. Commonly, therefore, this distinction is rather easy.
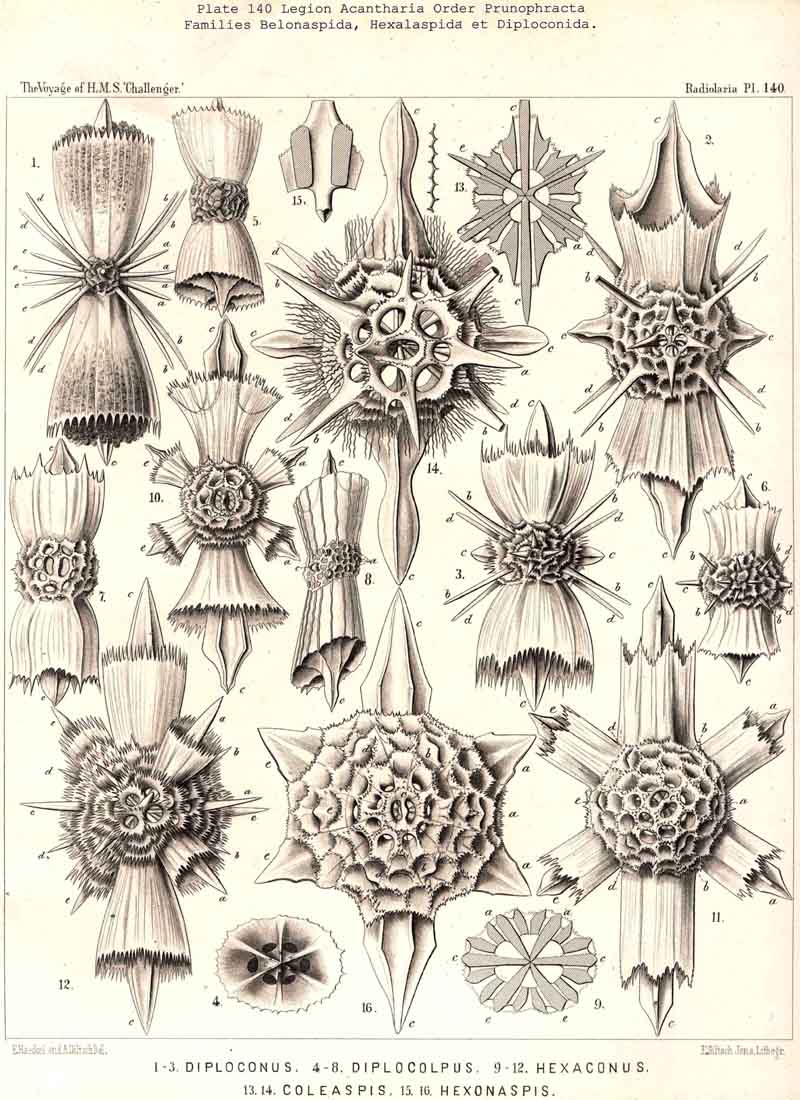
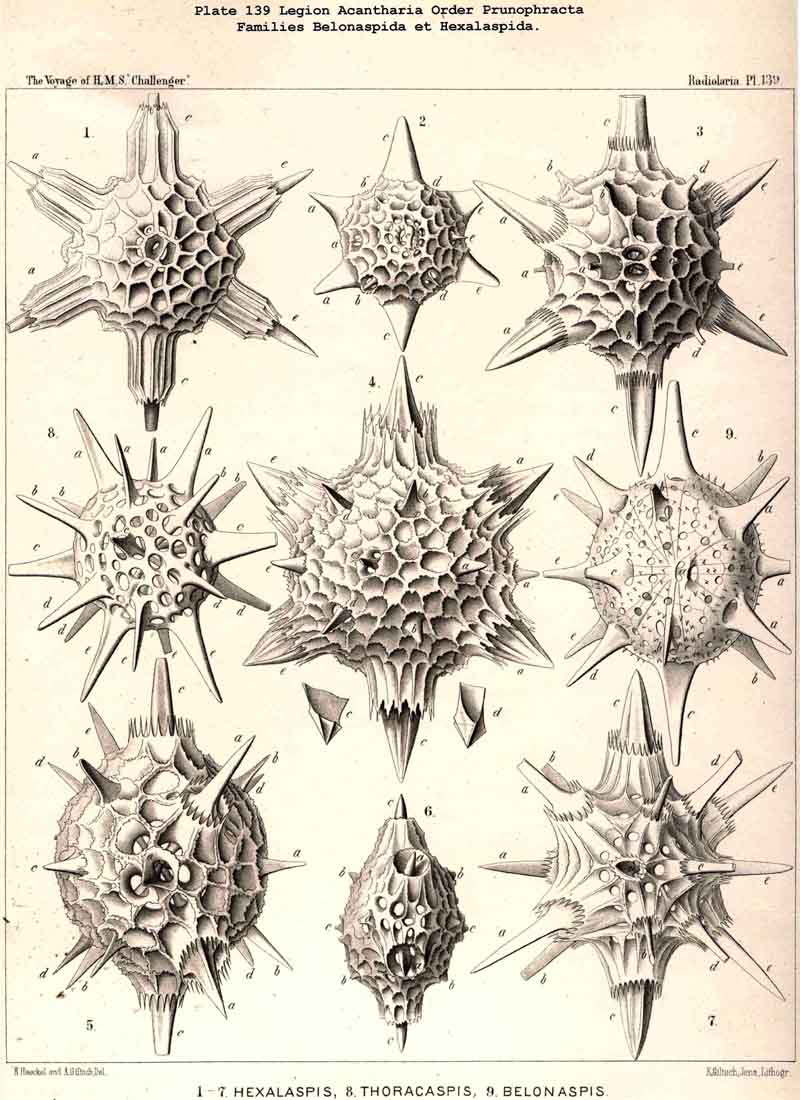
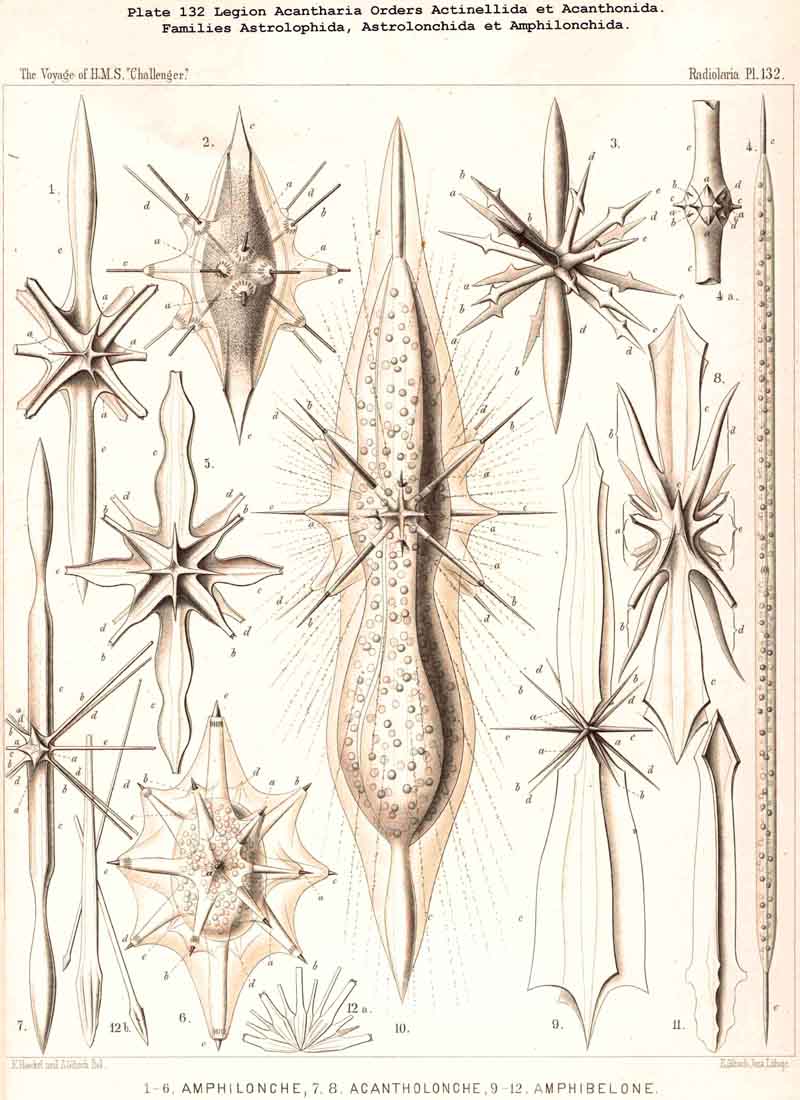
In the majority of the Icosacantha all four equatorial spines are exactly of the same form and size. But in four families the two opposite spines of one equatorial axis are much larger, or of another form, than those of the crossing axis. This is the case in the Amphilonchida, Belonaspida, Hexalaspida, and Diploconida. Therefore we here call the major equatorial axis (with larger spines) the "hydrotomical axis," and the minor axis (with smaller spines) the "geotomical axis." Correspondingly, the meridian plane, in which the two larger equatorial spines are placed (c1, c3) and the appertaining four polar spines (a1, a3, e1, e3) may be called the "hydrotomical plane "; in the remarkable family of Hexalaspida (Pl. 139) all six spines of this hydrotomical phane are much larger than the other fourteen. Perpendicular to this plane is the second perradial meridian plane, which we call the "geotomical plane"; in it lie the two smaller equatorial spines (c2,c4) and the corresponding four polar spines (a2, a4, e2, e4). In some Hexalaspida (Hexonaspis and Hexacolpus) the six spines of the hydrotomical plane become so preponderant that the other fourteen spines appear rudimentary; and in some of them the two equatorial spines of the hydrotomical plane are much larger than the four polar spines of the same plane. This curious relation reaches its maximum in the Diploconida (Pl. 140).
The different development of the two equatorial axes (of the larger hydrotomical and the smaller geotomical axis) is the first and most important cause of the peculiar forms, which are produced in the four cited families. We derive these terms also from the metaphor of the terrestrial globe. The hydrotomical plane is that meridian plane of the globe which intersects almost only the water-hemisphere (the island of Ferro in the Atlantic, the island of Pandora in the Pacific). Perpendicular to this is the geotomical plane, the meridian of which intersects great land-masses in both hemispheres (Bombay in India, Athabasca in Canada). Both poles of the smaller geotomical axis are everywhere equal (the East Indian and the Western American). However, both poles of the larger hydrotomical axis (the eastern Atlantic and the western Pacific) are in some genera very different, e.g., in Amphibelone among the Amphihonchida, and in Zygostaurus among the Quadrilonchida. In this case we call the anterior (commonly more developed) pole of the hydrotomical axis the frontal pole, the opposite posterior (commonly smaller) the caudal pole (Pl. 131, figs. 7, 8; Pl. 132, figs. 9, 10). On both sides of these (right and left) lie symmetrically the two equal poles of the geotomical lateral axis.
The promorphohogy of the ACANTHARIA demonstrates that the geometrical fundamental form in those groups is different. In the majority of the ACANTHARIA, where the two equatorial axes are equal, that form is a double square-pyramid or a "quadrate octahedron"; the four equal equatorial spines indicate the two diagonals of the square, which is the common base of the united regular four-sided pyramids ; their common axis is the spineless axis of the body; the ends of the polar spines fall on the edges of the pyramids, while the ends of the tropical spines fall on the halving lines of their faces. However, in those ACANTHARIA in which the two equatorial axes become different, the square double pyramid becomes changed into a rhombic double pyramid; the common base of the united pyramids is thus a rhombus; the hydrotomical axis is the larger, the geotomical axis the smaller diagonal of the rhombus.
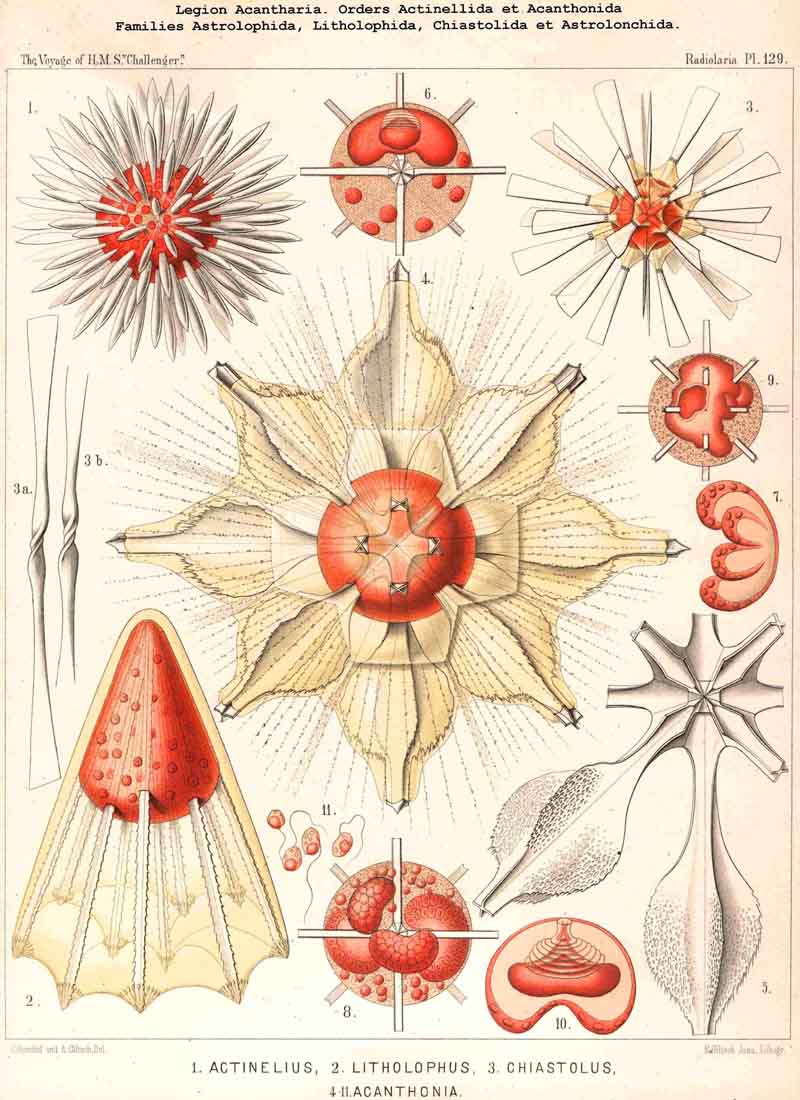
Opposed to the Icosacantha, under the name "Adelacantha," is the small group of Actinelida, in which the number and disposition of the radial spines is variable, not determined by the Müllerian law. Probably this group is the common ancestral stock, from which the Icosacantha have been derived by gradual development of their peculiar disposition. Probably the oldest and most primitive form of all ACANTHARIA is Actinelius, in which a variable and undetermined (often very large) number of radial spines is united in one common central point, and therefore forms a needle-sphere. Whilst here all spines (often more than a hundred) are of equal size and form, in the nearly allied Astrolophus large and small spines are intermingled. Both genera together form the small ancestral family of Astrolophida. In the strange family of Litholophida the radiah spines do not radiate within a spherical space (equally disposed in all directions), but within a quadrant or even an octant, forming a conical brush or pencil.
One very remarkable form of Actinelida is Actinastrum, forming the transition from these Adelacantha to the common regular Icosacantha. In the two observed species of Actinastrum we find thirty-two radial spines, twenty of which are disposed after the Müllerian law, as in the Icosacantha. The other twelve are four interradial equatorial spines (lying in the two secondary meridian planes) and eight perradial tropical spines (lying in the two primary meridian planes). Therefore here in each primary meridian plane are placed ten spines (two equatorial, four tropical, and four polar spines), whereas in each secondary meridian plane are placed six spines (two equatorial and four tropical). But here also all thirty-two spines are so regularly placed that their free distal ends fall into five parallel zones, four in each polar zone , eight in each tropical zone, and eight in the equatorial zone.
The Central Junction of the radial spines in the ACANTHARIA becomes effected in four different ways :-( 1) by simple apposition of the pyramidal central ends or bases; (2) by a basal leaf-cross, or by broad wings, four on each spine, supported one upon the other; (3) by a central concrescence of the meeting bases of all the twenty spines, growing perfectly together; and (4) by a concrescence in pairs of every two opposite spines. The most common and probably the original mode of junction is the firstby pyramidal apposition; the spines at the central base are pointed in the form of a pyramid, and the triangular faces of the neighbouring pyramids are simply placed upon one another. Often the small basal pyramids are imperfectly separated from the spines by an annular constriction. Commonly the basal pyramids of the four equatorial spines are six-sided, those of the sixteen other spines five-sided.
The second mode of junction, by a basal leaf-cross, is developed from the first and appears as a strengthening or a mechanical elaboration of it. Immediately above the basal pyramid arise from its radial edges four thin and broad triangular leaves or wings, and the meeting edges of the neighbouring wings are in apposition one with the other, so that between the bases of every three or four neighbouring spines a hollow pyramidal space remains open. The apex of such a pyramidal space is directed towards the centre of the body, but separated from it by the small basal pyramid; its open base is directed outwards. The twenty-two hollow pyramidal spaces are disposed regularly in four different groups :-(A) Four equatorial spaces, four-sided, each limited by two equatorial and two tropical spines (one canceral and one capricornal); (B) eight perizonal spaces (four northern and four southern), four-sided, each limited by one equatorial, two tropical, and one polar spine; (C) eight peripolar spaces (four northern and four southern), three-sided, each limited by one tropical and two polar spines; (D) two polar spaces (one northern and one southern), four-sided, each limited by four neighbouring polar spines.
The third mode of junction, by central concrescence of all twenty spines, was formerly regarded by me as an important peculiarity, sufficient for the separation of subfamiies and genera (Monogr. d. Radiol., 1862, pp. 399, 401 ; Prodromus, 1881, p. 466). But I found afterwards that in many species where the twenty spines commonly remain separated, accidentally they grow perfectly together and form one single piece of acanthina starrulet with twenty rays. Therefore I now think it is more natural to divide those species only into different subgenera.
A fourth and a very different mode of junction, quite sufficient for the distinction of different families, is the concrescence in pairs of every two opposite spines, lying in one diameter (in Acanthochiasma and Chiastolus). Here we obtain a number of "diametral spines" (each composed of two originally opposed radial spines) and all these diametral spines are crossed loosely near the central point of the body without any solid and permanent apposition (Chiastolida). However, in some species of this peculiar family the central part of the diametral spines is twisted like a screw or spirally convoluted (Pl. 129, figs. 2, 3).
The Form of the Radial Spines in the ACANTHARIA is extremely varied, and constitutes the main characters for the distinction of nearly four hundred species. But all these different forms may be reduced phylogenetically to three different fundamental forms :-(a) the cylindrical (with circular transverse section), (b) the two-edged (with elliptical or hanceolate transverse section), and (c) the four-edged (with square transverse section). No doubt the first (a) is the original primitive form, from which the two others are secondarily derived. Triangular spines never occur in the ACANTHARIA, whilst, however, they are common in the Sphaerellaria. The first and original form, the cylindrical spine, is either a true cylinder of equal thickness in its whole length , or it is more or less conical. Rarely the spine is in the distal half spindle-shaped, and thicker than in the basal half. The second form, the two-edged spine, is more or less compressed from two opposite sides; its two edges are either more blunt, rounded, or more acute, sharp; its transverse section in the former case is elliptical, in the latter case lanceolate or rhomboidal. Sometimes the two edges are broader and in the form of two thin opposite wings. The two-edged spines may be occasionally shorter, triangular or lanceolate, at other times longer sword-shaped or linear. The third form, the four-edged spine, has constantly a square transverse section; the sides of this square are either even or concave; in the latter case the four edges are broadened and wing-like, but in the former case not. The quadrangular spines are either prismatic (of equal breadth throughout their whole length) or pyramidal (becoming gradually thinner towards the distal apex).
The Apex of the Radial Spines, or their free distal end, is in the majority of ACANTHARIA simple, conical. In the minority it is either truncated or bifid, or four-sided pyramidal, often with two, rarely with four prominent parallel teeth. In some forms the bifid spines are so deeply cleft that they become forked. Much more interesting and more varied than these different forms of the distal end are those of the apophyses of the radial spines.
The Apophyses of the Radial Spines, or their "lateral transverse processes," are of the greatest importance for the morphological development of the whole subclass. Only in sixteen among the sixty-five genera of ACANTHARIA are the apophyses perfectly wanting; in the other genera they determine in the first place their general character. In the Acanthometra the apophyses remain perfectly free, whilst in the Acanthophracta their meeting ends or branches compose the latticed shell. All differences in form and shape of the apophyses can be reduced to only two primary modes; either the spine bears two opposite or four crossed apophyses; correspondingly all Acantharia apophysaria may be divided into two different main groups, the Zygapophysica (with two opposite lateral processes) and the Staurapophysica (with four crossed lateral processes opposite in pairs). Both groups have probably no direct phylogenetic connection, but seem to be derived independently from different stocks, and produce different families. The Zygapophysica are probably derived from Astrolonchida with two-edged spines (Zygacantha), and from this group arise the Diporaspida, the ancestral group of the majority of Acanthophracta. On the other hand the Staurapophysica are probably derived from Astrolonchida with four-edged spines (Acanthonia), and from this group arise the Tessaraspida. The apophyses of the Acanthonida are partly simple, partly branched or even latticed; the apophyses of the Acanthophracta are never simple, constantly branched and commonly latticed.
The Malacoma (or the whole soft body of the ACANTHARIA as opposed to the skeleton) exhibits some peculiarities which distinguish them from the other Radiolaria, as well in the structure of the central capsule and its nucleus as in that of the enveloping extra capsular body and the pseudopodia.
The Central Capsule is constantly spherical in the far greater number of the ACANTHARIA, viz. , in the following six families :- Astrolophida, Chiastolida, Astrohonchida, Dorataspida Sphaerocapsida, and Phractopeltida. Among these six families the Astrohonchida and Dorataspida are far greater and far richer in different forms than all the other families. The central capsule becomes ellipsoidal or cylindrical, prolonged in one axis, in the three families, Amphilonchida, Belonapsida, and Diploconida; it becomes discoidal or lenticular, by the shortening of one axis, in two families, viz., in the Quadrilonchida and Hexalaspida. Finally, the peculiar family Litholophida is distinguished by the conical form of its central capsule.
The Membrane of the central capsule in all ACANTHARIA is simple, commonly thin, sometimes very delicate; in some species it seems to be developed late, just immediately before the formation of the spores; but in no species is it completely missing. The membrane is constantly pierced by innumerable fine pores, for the emission of the pseudopodia; but in many species (and probably more or less in all ACANTHARIA) there is recognisable a certain regularity in the disposition of the numerous pseudopodia and of the pores by which they radiate from the capsule. Sometimes these pores are disposed in a regular network of ramified lines, whilst the meshes of this network are devoid of pores; in other cases they form regular tufts or bushes between the radial spines. Probably in no ACANTHARIA are the pores of the capsule membrane so numerous and so equally distributed throughout as in the SPUMELLARIA; we may therefore call the former ACTIPYLEA (in opposition to the latter as PERIPYLEA).
The Nucleus of the ACANTHARIA is constantly excentric, whilst it is originally constantly central in the SPUMELLARIA. This excentric position is a necessary consequence of the centrogenous development of the radial spines. Probably connected with this peculiarity is the other, that the nucleus assumes a peculiar, complicated structure, and that in the greater number of ACANTHARIA it becomes cleft very early, and that this cleavage is effected by a peculiar kind of gemmation, first detected and very accurately described by R. Hertwig (compare his Organismus d. Radiol., 1879, pp. 10-24). However, in the young ACANTHARIA the nucleus is constantly simple, and in a certain number of species its cleavage takes place late (as in the greater number of SPUMELLARIA).
The Endoplasm, or the intracapsular sarcode, exhibits in the greater number of ACANTHARIA a more or less distinct radial arrangement; but this is often concealed by the different enclosed products of the endoplasm oil-globules, vacuoles, red or different coloured pigment-granules, crystals, &c. Often it encloses a variable number of "yellow cells" (becoming green by mineral acids) to be considered as symbiotic xanthellae.
The Calymma or the jelly-veil, including the central capsule, in the ACANTHARIA is more or less voluminous, and commonly envelops the skeleton perfectly. In its surface is sometimes developed a peculiar network of "supporting fibres." A very peculiar product are the remarkable "Myophrisca' of the Acanthometra, which are wanting in the Acanthophracta; they were first detected by Johannes Müller, and figured as "Cilien-Kränze," afterwards explained by Hertwig as "contractile Fäden," siniilar to muscular fibrillae (compare below).
The Matrix, placed between the calymma and central capsule, in the majority of the ACANTHARIA is a rather thin layer of granular exoplasm.
The Pseudopodia arising from it are not so numerous as in the SPUMELLARIA, and not so equally disposed over the whole surface. Also their tendency to ramify, anastomose, and form networks seems to be much less developed. Commonly they are simple or little ramified. In many cases (and perhaps everywhere) there may be distinguished two different kinds of pseudopodia :-(l) Axopodia, or permanent pseudopodia (with axial filaments?), piercing the wall of the central capsule, and arising from the central mass of endoplasm; and (2) Collopodia, or variable pseudopodia (without axial filaments), arising outside the capsule from the matrix of extracapsular sarcode or from the exoplasm on the surface of the calymma. These and other differentiations seem to indicate that the pseudopodia in the ACANTHARIA are more highly developed than in the SPUMELLARIA, and justify the denomination of the former as "Actipylea."
Synopsis of the Orders and Suborders of ACANTHARIA.
|
|